



Introduction
Among the Neotropical dactylogyrids, members of Urocleidoides Mizelle & Price, Reference Mizelle and Price1964 stand out for their wide distribution in host groups, with species parasitizing the gills and nasal cavities of fishes of the orders Characiformes, Gymnotiformes, Cyprinodontiformes, and Siluriformes (Oliveira et al. Reference Oliveira, Santos-Neto, Tavares-Dias and Domingues2020; Zago et al. Reference Zago, Yamada, Yamada, Franceschini, Bongiovani and da Silva2020; Freitas et al. Reference Freitas, Bezerra, Meneses, Justo, Viana and Cohen2021; Oliveira et al. Reference Oliveira, da Silva, Vieira and Acosta2021). The genus Urocleidoides was proposed by Mizelle & Price (Reference Mizelle and Price1964) to accommodate U. reticulatus Mizelle & Price, Reference Mizelle and Price1964, collected from the gills of Poecilia reticulata Peters (Cyprinodontiformes, Poeciliidae) of the Capital Aquarium, Sacramento, California, USA. The genus was characterized by the presence of a sinistral vagina and copulatory complex comprising an accessory piece and a non-articulated male copulatory organ (MCO). Subsequently, Mizelle et al. (Reference Mizelle, Kritsky and Crane1968) and Mizelle & Kritsky (Reference Mizelle and Kritsky1969) reviewed the diagnosis of Urocleidoides and proposed new species from hosts of the orders Atheriniformes, Characiformes, Cypriniformes, Perciformes, and Siluriformes. However, Kritsky & Thatcher (Reference Kritsky and Thatcher1983) viewed the diversity of morphological structures of internal organ systems and absence of shared characteristics among previously known species of Urocleidoides as strongly suggesting the non-monophyly of the genus. Kritsky et al. (Reference Kritsky, Thatcher and Boeger1986) amended the diagnosis of the genus to only include monogenoids with the following main characteristics: presence of hook-shaped vaginal sclerite, coiled MCO with counterclockwise rings, and hook pairs 1 and 5 usually reduced.
Prior to the present study, from the species of Urocleidoides previously considered incertae sedis by Kritsky et al. (Reference Kritsky, Thatcher and Boeger1986), 14 have been synonymized or relocated into other genera as follows: Palombitrema Price & Bussing, Reference Price and Bussing1968 (1 species); Demidospermus Suriano, 1983 (1 species); Philocorydoras Suriano, 1986 (2 species); Sciadicleithrum Kritsky, Thatcher & Boeger, Reference Kritsky, Thatcher and Boeger1989 (1 species); Ameloblastella Kritsky, Mendoza-Franco & Scholz, Reference Kritsky, Mendoza-Franco and Scholz2000 (2 species), Aphanoblastella Kritsky, Mendoza-Franco & Scholz, Reference Kritsky, Mendoza-Franco and Scholz2000 (2 species), Diaphorocleidus Jogunoori, Kritsky & Venkatanarasaiah, Reference Jogunoori, Kritsky and Venkaranarasaiah2004 (3 species), Characithecium Mendoza-Franco, Reina & Torchin, Reference Mendoza-Franco, Reina and Torchin2009 (1 species); Nanayella Acosta et al., Reference Acosta, Mendoza-Palmero, da Silva and Scholz2019 (1 species) (Price & Bussing Reference Price and Bussing1968; Kritsky et al. Reference Kritsky, Thatcher and Boeger1989; Kritsky et al. Reference Kritsky, Mendoza-Franco and Scholz2000; Mendoza-Franco et al. Reference Mendoza-Franco, Posel and Dumailo2003; Jogunoori et al. Reference Jogunoori, Kritsky and Venkaranarasaiah2004; Mendoza-Franco et al. Reference Mendoza-Franco, Reina and Torchin2009; Mendoza-Palmero & Scholz Reference Mendoza-Palmero and Scholz2011; Yamada et al. Reference Yamada, Brandão, POF and da Silva2015; Acosta et al. Reference Acosta, Mendoza-Palmero, da Silva and Scholz2019). However, even though the correct taxonomic status of several species has been established, nine are still considered incertae sedis – namely, Urocleidoides stictus Mizelle, Kritsky & Crane, Reference Mizelle, Kritsky and Crane1968, Urocleidoides strombicirrus (Price & Bussing, 1967) Kritsky & Thatcher, Reference Kritsky and Thatcher1974, and Urocleidoides trinidadensis Molnar, Hanek & Fernando, 1974 reported from the gills of Characiformes; Urocleidoides gymnotus, Urocleidoides carapus, Urocleidoides virescens Mizelle, Kritsky & Crane, Reference Mizelle, Kritsky and Crane1968, and Urocleidoides advenai found in the gills of Gymnotiformes; and Urocleidoides amazonensis Mizelle & Kritsky, Reference Mizelle and Kritsky1969 and Urocleidoides catus Mizelle & Kritsky, Reference Mizelle and Kritsky1969 from the gills of Siluriformes.
The integration of morphological and molecular data has been used to improve the understanding of the taxonomic status of species of Urocleidoides, as well as to delimit the diagnosis of the genus. Gasques et al. (Reference Gasques, Graça, Prioli, Takemoto and Prioli2016) investigated the COI sequences of U. malabaricusi Rosim, Mendoza-Franco & Luque, Reference Rosim, Mendoza-Franco and Luque2011 and U. cuiabai Rosim, Mendoza-Franco & Luque, Reference Rosim, Mendoza-Franco and Luque2011 and detected that the mean divergence rate among the specimens of U. malabaricusi indicates the existence of the cryptic species. Acosta et al. (Reference Acosta, Mendoza-Palmero, da Silva and Scholz2019) used combined morphological and molecular data from the partial 28S rDNA gene and relocated Urocleidoides megorchis Mizelle & Kritsky, Reference Mizelle and Kritsky1969 reported in Surubim lima (Bloch & Schneider) (Siluriformes) to the genus Nanayella as Nanayella megorchis (Mizelle & Kritsky, Reference Mizelle and Kritsky1969) Acosta et al., Reference Acosta, Mendoza-Palmero, da Silva and Scholz2019. Furthermore, Zago et al. (Reference Zago, Yamada, Yamada, Franceschini, Bongiovani and da Silva2020) proposed a study based on morphological and molecular data (i. e., 28S rDNA and Cytochrome Oxidase I – COI) for species of Urocleidoides described from characiform and gymnotiform fishes. In their study, the COI mtDNA gene revealed a close relationship between U. strombicirrus (incertae sedis) and the other species of Urocleidoides (sensu stricto). These authors suggested that future investigation should focus on molecular characterization of stricto sensu and incertae sedis species to test the monophyly of Urocleidoides. The study of Oliveira et al. (Reference Oliveira, da Silva, Vieira and Acosta2021), also based on 28S rDNA analysis, supported the close relationship among some species of Urocleidoides and Cacatuocotyle papilionis Zago et al., Reference Zago, Franceschini, Müller and da Silva2018, suggesting that Urocleidoides may represent a non-monophyletic group.
The present study describes four new species of Urocleidoides reported from Erythrinidae (Characiformes) and Hypopomidae (Gymnotiformes) fishes from different hydrographic basins of the Northeastern Pará mesoregion (Eastern Amazon) based on morphological and molecular data (28S rDNA and COI mtDNA). In addition, it seeks to understand the phylogenetic relationships between some incertae sedis species (i.e., U. carapus and U. gymnotus) with species of Urocleidoides (sensu stricto) and other Neotropical dactylogyrids.
Material and methods
Host collection
Hosts were collected with the use of trammel net and landing nets in four locations in the Northeastern Pará mesoregion: Igarapé Maratininga – Moju River, municipality of Tailândia (2°27'55.7"S, 48°53'27.6"W); Balneário Aracu – Guamá River, municipality of Ourém (1°34'1.02"S, 47°9'52.35"W); Vila Perseverança – Palheta River, municipality of São Domingos do Capim (1°51'41.8"S, 47°38'26.5"W); and Vila Segredo – Segredo River, municipality of Capanema (1°5'32.44"S, 47°5'37.02"W).
Parasitological procedures
The gill arches were removed and placed in labeled vials containing heated water (~65°C). Each vial was shaken vigorously, and the sediment and gills were then fixed in 5% formalin for morphological studies or 96% ethanol for molecular characterization. In the laboratory, the content of each vial was examined with a stereoscopic microscope (LEICA S6D, Leica Microsystems, Wetzlar, Germany); the helminths found were removed from the gills or sediments using dissection needles and sent for morphological/molecular analysis. Monogenoid specimens intended for studies of internal structures were stained with Gomori’s Trichrome (Humason Reference Humason1979; Boeger & Vianna Reference Boeger, Vianna and Thatcher2006) and mounted in Dammar gum. For the study of sclerotized structures, the remaining specimens were mounted in Hoyer’s or Grey & Wess medium (Humason Reference Humason1979; Boeger & Vianna Reference Boeger, Vianna and Thatcher2006). The measurements were obtained according to the procedures of Mizelle & Klucka (Reference Mizelle and Klucka1953) and are presented in micrometers. The internal organs and other structures were measured in the dorsoventral view with an ocular micrometer. The length of curved or bent structures (i. e., anchors, bars, accessory piece) reflects the straight-line distances between ends. The total length of the MCO was obtained using ImageJ 1.43m (Rasband Reference Rasband2016). Hooks were classified according to Mizelle and Price (Reference Mizelle and Price1963). The averages of the measurements were calculated from the minimum and maximum length and the number of structures measured (n). Illustrations of the species and their structures were prepared using a drawing tube on a microscope with differential interference contrast and phase-contrast optics (LEICA DM 2500, Leica Microsystems, Wetzlar, Germany). Definitions of prevalence and mean intensity were calculated according to Bush et al. (Reference Bush, Lafferty, Lotz and Shostak1997). Type specimens, vouchers, and hologenophores presented in this study were deposited in the Helminthological Collection of the Instituto Oswaldo Cruz (CHIOC, Portuguese acronym), Rio de Janeiro, Rio de Janeiro State, Brazil. Acting in accordance with the regulations in article 8.5 of the amended 2012 version of the International Code of Zoological Nomenclature, details of the new taxa have been submitted to ZooBank.
DNA extraction, amplification, and sequencing
Each parasite specimen submitted to molecular analysis was divided with small dissection needles using a stereoscopic microscope for morphological identification. When the species was identified using haptoral structures, the anterior region of the body was placed in a 1.5 ml microtube with 96% ethanol for DNA extraction. However, when the morphology of the MCO was used for identification, the posterior region of the body was used for DNA extraction. The anterior or posterior regions of the parasite body were mounted in Hoyer medium between the slide and cover slip to identify the species. Genomic DNA was extracted with the DNeasy® Blood and Tissue kit (QIAGEN, Hilden, Germany), according to the manufacturer’s protocol, with a final volume of 30 μl. DNA concentration was verified using a NanoDrop 2000 spectrophotometer (Thermo Fisher Scientific, Massachusetts, USA).
The partial region of the 28S rDNA gene was amplified by PCR in two steps. In the first step, DNA was amplified with primer pairs 1200F (Littlewood & Olson Reference Littlewood, Olson, Littlewood and Bray2001) and D2 (Wu et al. Reference Wu, Zhu, Xie and Li2006). In the second step, nested PCR was performed using C1 (Wu et al. Reference Wu, Zhu, Xie and Li2006) and D2 primers, amplifying a fragment of ~800 bp. The amplification program was configured for an initial denaturation step of 94°C for 5 minutes, followed by 35 cycles of 94°C for 45 seconds, 50°C for 30 seconds, 72°C for 90 seconds, and a final extension of 72°C for 7 minutes. Nested PCR was conducted with 1 μl of the PCR product, diluted 1:1 in ultrapure water, with the same amplification program described above. Sequencing was performed using C1 and D2 primers. The partial sequence of gene COI mtDNA was amplified using the primers COI_Mono_5 and COI_Mono_3 and/or COI_Mono_int3 (Plaisance et al. Reference Plaisance, Rousset, Morand and Littlewood2008). The amplification program was configured for an initial denaturation step of 94°C for 3 minutes, followed by 40 amplification cycles at 94°C for 30 seconds, 44°C for 30 seconds, 72°C for 4 minutes, and a final extension of 72°C for 7 minutes (Plaisance et al. Reference Plaisance, Rousset, Morand and Littlewood2008). COI_Mono_int3 was used for sequencing.
PCRs were performed in a Matercycler® Nexus (Eppendorf, Hamburg, Germany) with a final volume of 25 μl using DreamTaq Green PCR Master Mix (2×) (Thermo Scientific Wilmington, USA), following the manufacturer’s recommendations. The reactions were performed with 0.5 mM of each primer and 3 μl of extracted DNA. PCR products were run on 2% agarose gel stained with GelRed (Biotium Inc., Hayward, California, USA), and DNA quality was assessed in an ultraviolet transilluminator. The amplified products were purified with QIAquick PCR Purification Kit (QIAGEN, Hilden, Germany). Sequencing was performed with Big Dye® Terminator Cycle Sequencing Kit v.3.1 (Applied Biosystems, California, USA) in an ABI 3500 XL automatic sequencer (Applied Biosystems, California, USA) at the Instituto de Estudos Costeiros (IECOS), Universidade Federal do Pará (UFPA), Pará, Brazil.
Phylogenetic analyses
Sequences of the partial 28S rDNA gene were obtained from eight species of Urocleidoides, along with six sequences from the partial COI mtDNA gene. The sequences obtained were submitted to BLAST analysis (http://blast.ncbi.nlm.nih.gov) to verify similarities with other monogenoid sequences. The partial 28S rDNA sequences obtained in the present study were aligned with 30 species of Dactylogyridae, and three species of Diplectanidae were used as an outgroup (Murraytrema pricei Bychowsky & Nagibina, Reference Bychowsky and Nagibina1977, Pseudorhabdosynochus lantauensis (Beverley-Burton & Suriano, 1981) Kritsky & Beverley-Burton, Reference Kritsky, Thatcher and Boeger1986, and Pseudorhabdosynochus epinepheli (Yamaguti, 1938) Kritsky & Beverley-Burton, Reference Kritsky, Thatcher and Boeger1986) retrieved from GenBank (Table 1). The sequences obtained for the partial COI mtDNA gene were aligned with 21 sequences from Urocleidoides, with one sequence from Acanthocotylidae (Acanthocotyle gurgesiella Ñacari, Sepulveda, Escribano & Oliva, Reference Ñacari, Sepulveda, Escribano and Oliva2017) used as an outgroup (Table 1).
Table 1. List of monogenoids included in the phylogenetic analyses with details of the parasite species, host species, host family, locality, and GenBank accession numbers

The sequences obtained in the present study are in bold. *Outgroup used
The sequences were aligned with the Clustal W algorithm (Thompson et al. Reference Thompson, Higgins and Gibson1994) implemented in Geneious version 7.1.3 (Kearse et al., Reference Kearse, Moir, Wilson, Stones-Havas, Cheung, Sturrock, Buxton, Cooper, Markowitz, Duran, Thierer, Ashton, Mentjies and Drummond2012). Genetic divergence was determined using the p-distance model matrix in MEGA X (Kumar et al. Reference Kumar, Stecher, Li, Knyaz and Tamura2018). The JModelTest 2.1.1 software (Posada Reference Posada2008) was used to select the most appropriate evolutionary model for the Maximum Likelihood (ML) and Bayesian Inference (BI) analyses based on the Akaike information criterion (AIC). The evolutionary model selected was GTR + I + G for the partial 28S rDNA and TPM3uf + I + G for the partial COI mtDNA. The search for the ML tree was performed with bootstrap confidence determined by performing 1,000 replicates using PhyML 3.0 implemented via the web server on the ATGC - Montpellier Bioinformatics Platform (http://www.atgcmontpellier.fr/phyml/) (Guindon et al. Reference Guindon, Dufayard, Lefort, Anisimova, Hordijk and Gascuel2010). BI analysis was performed using MrBayes v.3.2 (Ronquist & Huelsenbeck Reference Ronquist and Huelsenbeck2003). The TPM3uf + I + G evolutionary model indicated by JModelTest is not implemented in MrBayes, so it was replaced with the closest over-parameterized model available (GTR + I + G). BI analysis was implemented with posterior probability estimated from 1 million generations with two independent runs of four Markov Chain Monte Carlo (MCMC) with algorithms sufficient to keep the average standard deviation below 0.001. Trees were sampled every 1,000th generation, with diagnostics every 1,000th generation and a burn-in period covering the first 25,000 generations. Tracer v. 1.6 (Rambaut et al. Reference Rambaut, Suchard, Xie and Drummond2014) was used to verify convergence and confirm the effective sample size (ESS) to provide reasonable estimates of the variance in model parameters (i. e., ESS values > 200). Only nodes with a posterior probability above 90% and bootstrap above 60% were considered. Phylogenetic trees were generated in FigTree v.1.4.3 (Rambaut Reference Rambaut2012) and edited using CorelDraw 2019©.
Results
Our present study provides information on the evolutionary relationships between species of Urocleidoides using phylogenetic analyses based on molecular data (28S rDNA and COI mtDNA). In addition, four new species are reported and described from Characiformes (Erytrinidae) and Gymnotiformes (Hypopomidae) hosts from the Eastern Amazon, expanding the genus to 52 valid species (Table 2).
Table 2. List of species Urocleidoides

CHA=Characiformes; CYP=Cyprinodontiformes; GYM= Gymnotiformes; SIL=Siluriformes.
Molecular data and phylogenetic inferences
Partial sequences of the 28S rDNA gene were obtained for four new species of Urocleidoides (U. atilaiamarinoi n. sp. – 767 bp long, U. vanini n. sp. – 773 bp long, U. macrosoma n. sp. – 731 bp long, and U. nataliapasternakae n. sp. 766 bp long), as well as for four previously described species (U. carapus [hologenophore, CHIOC No. 40202] – 750 bp long, U. gymnotus [hologenophore, CHIOC No. 40203] – 741 bp long, Urocleidoides naris Rosim, Mendoza-Franco & Luque, Reference Rosim, Mendoza-Franco and Luque2011 [hologenophore, CHIOC No. 40205] – 762 bp long, and Urocleidoides brasiliensis Rosim, Mendoza-Franco & Luque, Reference Rosim, Mendoza-Franco and Luque2011 [hologenophore, CHIOC No. 40201] – 731 bp long).
After trimming the ends, the aligned 28S rDNA sequences had a length of 660 bp. ML and BI analyses based on the partial 28S rDNA gene recovered similar tree topologies but differed in the posterior probability (P) and bootstrap (B) values. The phylogenetic analyses revealed a tree with two major clades, Clades A and B (Figure 1). Clade A is well-supported and divided into two subclades (Clades A1 and A2), which group species of monogenoid parasites of Siluriformes. Clade A1 groups the species that have been reported as parasitizing pimelodids and doradids, whereas its sister group, Clade A2, comprises monogenoids from loricariids and heptapterids.

Figure 1. Molecular phylogeny of Dactylogyridae estimated by Bayesian and Maximum Likelihood analyses inferred using the partial 28S rDNA gene (alignment length of 660 bp). The new species sequenced in the present study are presented in bold. The sequences of the other species were retrieved from GenBank. The bootstrap (ML) and posterior probability (BI) supports are presented between branches (values of posterior probability < 0.90 and bootstrap < 60 are not shown). The length of the scale bar indicates the substitution numbers per site.
Clade B is divided into two subclades (Clades B1 and B2) (Figure 1). Clade B1 comprises Urocleidoides spp. (Characiformes and Gymnotiformes), Cacatuocotyle papilionis (Characiformes: Characidae), Heteropriapulus spp., Trinigyrus anthus Franceschini, Acosta et al., Reference Franceschini, Acosta, Zago, Müller and da Silva2020, Unilatus unilatus Mizelle & Kritsky, Reference Mizelle and Kritsky1967 (Siluriformes: Loricariidae), and Mymarothecium viatorum Boeger, Piasecki, Sobecka, Reference Boeger, Piasecki and Sobecka2002 (Characiformes: Serrasalmidae). The clade formed by the species of Urocleidoides showed significant support for both analyses (BI, P = 1 and ML, B = 82). One group consists of the species that parasitize anostomids (U. paradoxus Kritsky, Thatcher & Boeger, Reference Kritsky, Thatcher and Boeger1986, U. sinus Zago et al., Reference Zago, Yamada, Yamada, Franceschini, Bongiovani and da Silva2020 and U. digitabulum Zago et al., Reference Zago, Yamada, Yamada, Franceschini, Bongiovani and da Silva2020) (BI, P = 1 and ML, B =100), parondontids (Urocleidoides tenuis Zago et al., Reference Zago, Yamada, Yamada, Franceschini, Bongiovani and da Silva2020; U. indianensis Oliveira et al., Reference Oliveira, da Silva, Vieira and Acosta2021 and U. parodoni Oliveira et al., Reference Oliveira, da Silva, Vieira and Acosta2021) (BI, P = 1 and ML, B = 100), and erythrinids (U. vanini n. sp. and U. atilaiamarinoi n. sp.) (BI, P = 1 and ML, B = 99). The other group is made up of species that parasitize Gymnotiformes (U. carapus, U. gymnotus, U. uncinus Zago et al., Reference Zago, Yamada, Yamada, Franceschini, Bongiovani and da Silva2020 [Gymnotidae], and U. nataliapasternakae n. sp. [Hypopomidae]) (BI, P = 1 and ML, B = 100) and H. malabaricus (Erythrinidae) (U. brasiliensis, U. naris, and U. macrosoma n. sp.) (BI, P = 1 and ML, B = 99). The group formed by C. papilionis and the reported species of Urocleidoides from Gymnotiformes and H. malabaricus shows low support for BI and ML. Clade B2 (BI, P = 1 and ML, B = 97) appears as a sister group to clade B1 and consists of species of monogenoids that parasitize pimelodids (Unibarra paranoplatensis Suriano & Incorvaia, 1995 and Ameloblastella edentesis Mendoza-Franco, Mendoza-Palmero & Scholz, Reference Mendoza-Franco, Mendoza-Palmero and Scholz2016), heptapterids (Ameloblastella chavarriai (Price, 1936) Kritsky, Mendoza-Franco & Scholz, Reference Kritsky, Mendoza-Franco and Scholz2000), and doradids (Vancleaveus janauacaensis Kritsky, Thatcher & Boeger, Reference Kritsky, Thatcher and Boeger1986) (Figure 1).
The partial sequences of the COI mtDNA gene were obtained for three new species of Urocleidoides (U. vanini n. sp. – 695 bp long, U. macrosoma n. sp. – 733 bp long, and U. nataliapasternakae n. sp. – 764 bp long), as well as for three previously described species (U. carapus [hologenophore, CHIOC No. 40202] – 717 bp long, U. gymnotus [hologenophore, CHIOC No. 40203] – 765 bp long, and U. naris [hologenophore, CHIOC No. 40205] – 679 bp long). The alignment obtained had a length of 347 bp, and both ML and BI analyses recovered similar tree topologies. Clade A of the phylogenetic tree is well-supported by BI analysis (P = 0.98) and divided into the clades A1 and A2 (Figure 2). Clade A1 groups the species that occur in Characiformes (Characidae and Parodontidae) and Gymnotiformes (Hypopomidae and Gymnotidae). Subclade A1’, which groups species of Urocleidoides found in Characiformes (Characidae) and Gymnotiformes (Hypopomidae and Gymnotidae), was poorly supported by both analyses (BI and ML). However, the subclade within A1’ comprised of the species that parasitize Gymnotiformes (Hypopomidae and Gymnotidae) is well-supported by BI analysis (P = 0.95), and groups Urocleidoides species that are both incertae sedis (U. gymnotus, U. carapus) and sensu stricto (U. cultellus, U. uncinus, and U. nataliapasternakae n. sp.). Urocleidoides tenuis, which parasitizes parodontids, appears as a sister species to species parasitizing Characiformes (Characidae) and Gymnotiformes (Hypopomidae and Gymnotidae). Clade A2, however, groups the species that occur in anostomids and erythrinids. In this clade, Urocleidoides digitabulum appears as a sister group of U. sinus and of the species that parasitize erythrinids. However, this group has low support and does not indicate such a relationship between these species. Likewise, the relationship between U. sinus and the species that occur in erythrinids is also not supported due to the low P and B values for both analyses. In contrast, the clade comprising the species that parasitize erythrinids is well-supported by the BI analysis (P = 0.98), but the support values between species are low.

Figure 2. Molecular phylogeny of Dactylogyridae estimated by Bayesian and Maximum Likelihood analyses inferred using the partial COI mtDNA gene (alignment length 347 bp). The new species sequenced in the present study are presented in bold. The sequences of the other species were retrieved from GenBank. The bootstrap (ML) and posterior probability (BI) supports are presented between branches (values of posterior probability < 0.90 and bootstrap < 60 are not shown). The length of the scale bar indicates the substitution numbers per site.
Genetic divergence of 28S rDNA was estimated only for monogenoid species belonging to Clade B1 (Table 3). The genetic divergence between Urocleidoides spp. and the other dactylogyrid species in Clade B1 ranges between 18.4 and 25.8% (203–349 bp) (Table 3). Urocleidoides spp. and Cacatuocotyle papilionis diverge at rates between 20.5 and 25% (208–322 bp); Urocleidoides spp. and Heteropriapulus spp. between 21.1 and 24.8% (205–328 bp); Urocleidoides spp. and Unilatus unilatus between 18.4 and 23% (203–315 bp); Urocleidoides spp. and Trinigyrus anthus between 20.3 and 25.2% (211–349 bp), and Urocleidoides spp. and Mymarothecium viatorum between 21.3 and 25.8% (212–244 bp).
Table 3. Pairwise genetic identities of 28S rDNA sequences selected from Dactylogyridae species of Clade B1

The upper triangular matrix shows the number of nucleotide differences, and the lower triangular matrix shows the differences in terms of percentage of nucleotides. Sequences obtained in the present work are in bold.
Among the species of Urocleidoides parasitizing Gymnotiformes (U. caparus, U. gymnotus, U. uncinus, and U. nataliapasternakae n. sp.), the genetic divergence ranges from 2 to 12.7% (32–154 bp). Among the species that parasitize anostomids (U. paradoxus, U. sinus, and U. digitabulum), the divergence varies between 7.8 and 11.3% (142–157 bp). For the species that parasitize H. malabaricus (Characiformes: Erythrinidae) (U. naris, U. brasiliensis, and U. macrosoma n. sp.), divergence ranges from 3.9 to 9% (43–87 bp). Urocleidoides atilaiamarinoi n. sp. reported for H. unitaeniatus (Agassiz) (Characiformes: Erithrinidae) diverges from the other species that parasitize H. malabaricus by between 19 and 21% (207–214 bp). Urocleidoides vanini n. sp. from Erythrinus erythrinus (Bloch & Schneider) (Characiformes: Erythrinidae) differs from the remaining species of Urocleidoides from H. malabaricus by between 20.7 and 21.5% (192–202 bp), and U. vanini n. sp. differs 8% (90 bp) from U. atilaiamarinoi n. sp. The species that parasitize characiform fish from the family Parodontidae (U. tenuis, U. indianensis, and U. parodoni) diverge from each other from 2.1 to 10.4%. (28–158 bp). For COI mtDNA, genetic divergence was estimated and compared for the species of Urocleidoides, varying between 14.7 and 30.8% (50–102 bp) (Table 4).
Table 4. Pairwise genetic identities of COI mtDNA sequences selected from Dactylogyridae species of Clade B1

The upper triangular matrix shows the number of nucleotide differences, and the lower triangular matrix shows the differences in terms of percentage of nucleotides. Sequences obtained in the present work are in bold.
Taxonomic summary
Class Monogenoidea Bychoswky, 1937
Order Dactylogyridea Bychoswky, 1937
Dactylogyridae Bychowsky, 1933
Urocleidoides Mizelle & Price, Reference Mizelle and Price1964
Urocleidoides vanini n. sp. (Figure 3)

Figure 3. Urocleidoides vanini n. sp. a. Holotype, whole body; b. copulatory complex; c. ventral bar; d. dorsal bar; e. hook pair 5; f. hook pair 1; g. hook pairs 2, 3, 4, 6, and 7; h. dorsal anchor; i. ventral anchor. Scales: a. 50μm scale, b–i. 25μm scale.
Type host. Erythrinus erythrinus (Bloch & Schneider)
Type locality. Vila Perseverança, Palheta River (Guamá River Basin), municipality of São Domingos do Capim, Pará, Brazil (1°51'41.8"S, 47°38'26.5"W).
Site of infestation. Gills.
Prevalence. 100% of 2 hosts examined.
Average intensity. 4.5 parasites per host.
Specimens deposited. Holotype (CHIOC No. 40208a); 9 paratypes (CHIOC No. 40208b–f, 40209a–d), 1 hologenophore (CHIOC No. 40208g).
Molecular sequence data. The partial 28S rDNA (773 bp) and COI mtDNA (695 bp) sequences obtained from one specimen (GenBank accession numbers OR270736 and OR285309, respectively).
Etymology. The specific name of the species is a tribute to zoologist Sergio Antonio Vanin (1948–2020), a lover of zoology who dedicated his life to the studies of systematics and taxonomy and contributed to the education of new generations of Brazilian zoologists.
Number of ZooBank. C053980D-B95E-4FC1-80F0-85932FF1B3CD.
Description. (Based on 10 adult specimens – 5 mounted on Gomori Trichrome, 5 mounted on Hoyer’s medium). Body elongated, robust, foliform, total length excluding haptor 157 (135–167; n=5), total width at level of germarium 112 (92–150; n=5) (Figure 3a). Cephalic lobes (4) moderately developed, 2 terminal and 2 bilateral; 3 pairs of head organs; cephalic glands not observed (Figure 3a). Eyes (2 pairs) equidistant; accessory chromatic granules distributed near pharynx (Figure 3a). Pharynx suboval, muscular, 21 (19–23; n=5) long, 16 (15–17; n=5) wide; esophagus short (Figure 3a). Genital pore midventral, anterior to copulatory complex. Genital atrium non-sclerotized. Gonads overlapping, testis dorsal to germarium (Figure 3a). Oviduct, Mehlis’ glands, uterus, egg, prostatic reservoir, seminal receptacle not observed. Testis oval 22 (19–24; n=3) long, 16 (14–17; n=3) wide. Copulatory complex comprising MCO, accessory piece. MCO sclerotized, tubular with three and a half counterclockwise rings, 194 (187–213; n=4) long, base with sclerotized cap; proximal portion of MCO slightly expanded, distal aperture acute. Accessory piece located in distal portion of MCO, not articulated with base of MCO, comprising an elongated sheath (Figure 3b). Seminal vesicle sigmoid (Figure 3a). Vaginal pore sinistral, ventro-marginal; vaginal vestibule broad, slightly sclerotized; vaginal canal muscular, sigmoid (Figure 3a). Vaginal sclerite absent. Germarium elongated 50 (42–60; n=5) long, 25 (22–27; n=5) wide. Vitellaria dense coextensive with gut, absent in regions of reproductive organs. Haptor hexagonal 38 (31–44; n=5) long, 76 (71–79; n=5) wide (Figure 3a). Anchors similar; each with well-developed superficial root, depressed at distal surface; short deep root, rounded at distal surface; shaft evenly curved, point; point extending past level of tip of superficial root; ventral anchor base 11 (11–12; n=5) long, inner 28 (25–33; n=5) long, outer 29 (27–31; n=5) long, (Figure 3i); dorsal anchor base 12 (11–13; n=5) long, inner 19 (18–20; n=5) long, outer 26 (24–26; n=5) long, (Figure 3h). Ventral bar 29 (25–35; n=7) long, open V-shaped with enlarged ends (Figure 3c); dorsal bar 28 (24–34; n=7) long, open U-shaped with rounded ends (Figure 3d). Hook pairs 2, 3, 4, 6, and 7 26 (24–29; n=6) long (Figure 3g) similar in morphology with shank divided into two subunits, thumb slightly depressed, slightly curved shaft, delicate point; hook pairs 1 and 5 reduced in size 13 (11–14; n=7) long; hook pair 1 with shank divided into two subunits, thumb erect, curved shaft, delicate point (Figure 3f); hook pair 5 with erected thumb, curved shaft, delicate point, lacking dilated shank portion (Figure 3e).
Remarks. The molecular results in the present study support the validity of U. vanini n. sp. as well as U. carapus and U. gymnotus as members of Urocleidoides (see Molecular data and phylogenetic inferences section). These three species differ from their cogeners by lacking the vaginal sclerite. The new species differs from U. carapus mainly by possessing a uniform dorsal anchor, whereas U. carapus has a dorsal anchor with point presenting ornamentation as sclerotized shredded filaments. Urocleidoides vanini n. sp. also can be distinguished from U. gymnotus mainly by having a MCO with three and a half rings (seven to nine rings in U. gymnotus).
Urocleidoides atilaiamarinoi n. sp. (Figure 4)
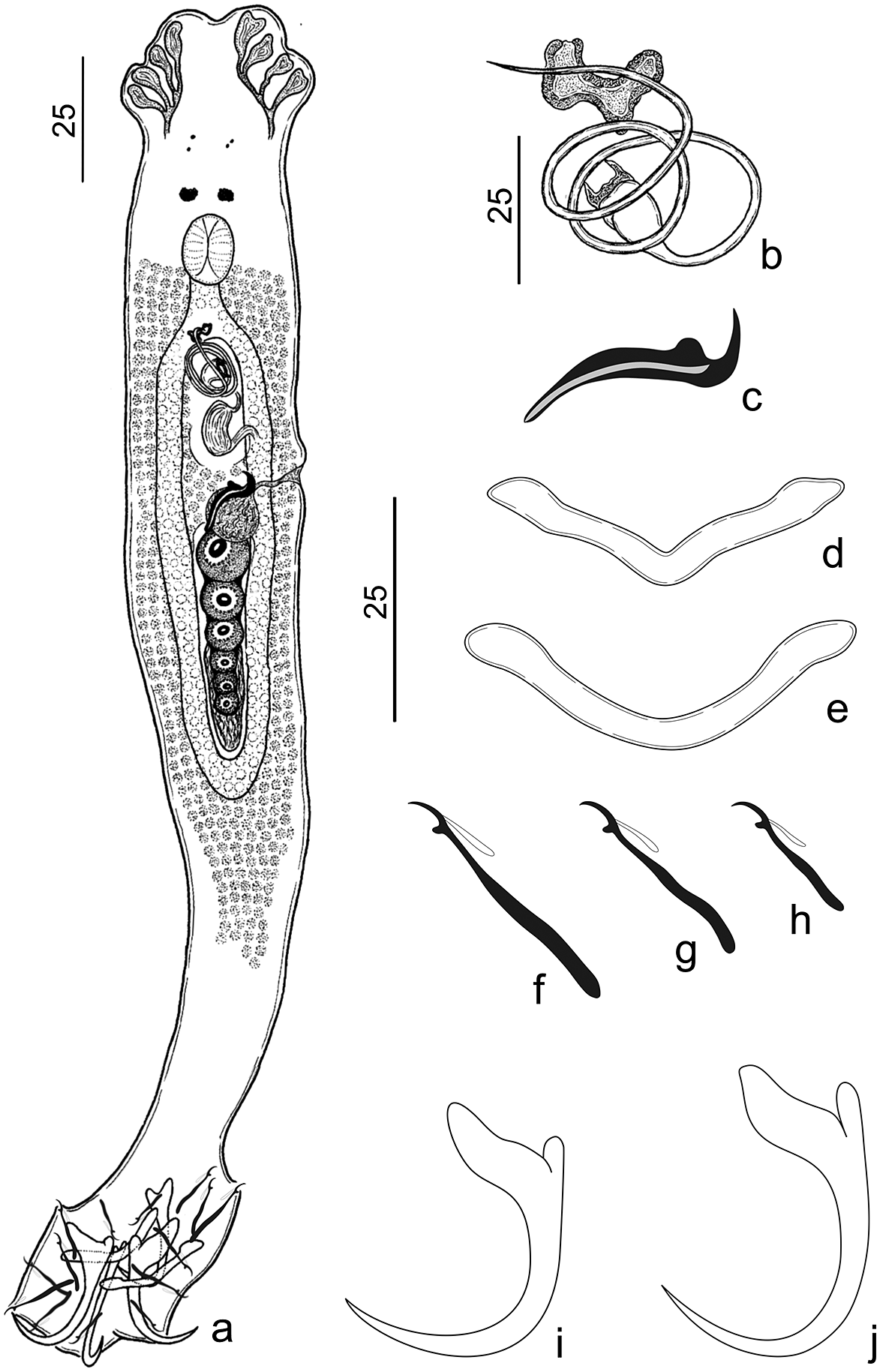
Figure 4. Urocleidoides atilaiamarinoi n. sp. a. Whole body – composite (ventral); b. copulatory complex; c. vaginal sclerite; d. ventral bar; e. dorsal bar; f. pair hook 7; g. hook pairs 2, 3, 4, and 6; h. hook pair 1; i. dorsal anchor; j. ventral anchor. Scales: a. 25μm scale, b–j. 25μm scale.
Type host. Hoplerythrinus unitaeniatus (Agassiz) (Characiformes: Erythinidae).
Type locality. Igarapé Maratininga (Moju River Basin), municipality of Tailândia, Pará, Brazil (02°27’55.7”S, 48°53’27.6”W).
Site of infestation. Gills.
Prevalence. 43% (3 of 7 hosts examined).
Average intensity. 5 parasites per host.
Other hosts and locations. Hoplerythrinus unitaeniatus (prevalence: 33% [1 of 3 hosts]; average intensity: 7 parasites per host), Balneário Aracu (Guamá River Basin), municipality of Ourém, Pará, Brazil (1°34’1.02”S, 47° 9’52.35”W).
Specimens deposited. Holotype (CHIOC No. 39995a); 14 paratypes (CHIOC No. 39995b–f, 39996a–c, 39997a–f), 1 hologenophore (CHIOC No. 40000); 7 vouchers (CHIOC No. 39998a–c, 39999a–d).
Molecular sequence data. The partial 28S rDNA (767 bp) sequence obtained from one specimen (GenBank accession number OR270164).
Etymology. The specific name of the species is a tribute to the biologist and science communicator Atila Iamarino, who is dedicated to studies related to genetics of microorganisms and works tirelessly to communicate scientific research to the general population.
Number of ZooBank. 8684EAB5-4587-44E1-9F25-9255CDE5FEE4.
Measurements. Table 5.
Table 5. Measurements (μm) of Urocleidoides atilaiamarinoi n. sp., gill parasite of Hoplerythrinus unitaeniatus from two locations

* Type-locality; MCO = male copulatory organ.
Description. (Based on 16 adult specimens – 2 mounted on Gomori’s Trichrome, 11 mounted on Hoyer’s medium, 3 mounted on Gray & Wess medium). Body elongated, fusiform, total length excluding haptor 209 (175–237; n=12), total width at level of germarium 75 (57–97; n=13) (Figure 4a). Cephalic lobes (4) well-developed, 2 terminal and 2 bilateral; 4 pairs of head organs; cephalic glands not observed (Figure 4a). Eyes (2 pairs) equidistant, posterior pair greater than anterior; anterior pair with few granules observed, accessory chromatic granules present or absent (Figure 4a). Pharynx oval, muscular 16 (15–18; n=8) long, 16 (12–17; n=8) wide; esophagus short (Figure 4a). Genital pore midventral, anterior to copulatory complex. Genital atrium non-sclerotized. Gonads overlapping, testis dorsal to germarium (Figure 4a). Oviduct, Mehlis’ glands, uterus, egg, prostatic reservoir not observed. Testis elongated 23 (n=1) long, 8 (n=1) wide. Copulatory complex comprising MCO, accessory piece. MCO sclerotized, tubular with two and a half counterclockwise rings, 120 (96–129; n=11) long, bulbous base with sclerotized cap, distal aperture acute (Figure 4b). Accessory piece located in distal portion of MCO, not articulated with base of MCO, dumbbell-shaped (Figure 4b). Vaginal pore ventro-marginal, vaginal vestibule slightly sclerotized, vaginal canal muscular, slightly sigmoid. Vaginal sclerite 30 (22–38; n=9) long, sickle-shaped with longitudinal superficial groove, thumb short, point curved, elongated (Figure 4c). Germarium elongated 37 (n=1) long, 10 (n=1) wide. Seminal receptacle spherical 12 (n=1) long, 11 (n=1) wide. Vitellaria dense coextensive with gut, absent in regions of reproductive organs. Haptor hexagonal 48 (32–55; n=12) long, 68 (47–90; n=12) wide (Figure 4a). Ventral anchor with elongate slightly depressed tip of superficial root; elongate deep root, rounded at distal surface; evenly curved shaft, point; point extending past level of tip of superficial root, base 12 (11–14; n=12) long, inner 26 (23–28; n=12) long, outer 24 (22–27; n=12) long (Figure 4j). Dorsal anchor with elongate superficial root; short deep root, rounded at distal surface; evenly curved shaft, point; point extending past level of tip of superficial root, base 10 (10–11; n=11), inner 23 (21–24; n=11) long, outer 22 (20–24; n=11) long (Figure 4i). Ventral bar 34 (29–40; n=12) long, open V-shaped, with ends slightly tapered (Figure 4d); dorsal bar 34 (28–40; n=12) long, open U-shaped with rounded ends (Figure 4e). Hook pairs similar in morphology with shank divided into two subunits, proximal dilation comprising 2/3 of shank length, thumb erect, elongated slightly curved shaft, delicate point, filament hook loop extended to near beginning of shank dilation. Hook pairs 1 and 5 reduced in size, pair 1 14 (12–16; n=12) long (Figure 4h); pair 5 15 (14–16; n=10) long; pairs 2, 3, 4, 6 22 (20–23; n=12) long (Figure 4g); pair 7 27 (25–29; n=13) long (Figure 4f).
Remarks. Urocleidoides atilaiamarinoi n. sp. resembles Urocleidoides bulbophallus Ferreira et al., Reference Ferreira, Rodrigues, Cunha and Domingues2017 since they share a MCO with bulbous base. However, the new species differs from U. bulbophallus due the numbers of MCO rings (2 ½ rings in U. atilaiamarinoi n. sp. and 1 ½ in U. bulbophallus) and the morphology of the accessory piece (dumbbell-shaped in U. atilaiamarinoi n. sp., and a bent sheath, ‘e’ shape in U. bulbophallus). Also, they differ on the comparative size of anchors and bars. In U. atilaiamarinoi n. sp., the anchors and bars are approximately similar in size, whereas U. bulbophallus has ventral anchors and ventral bar twice as large as dorsal anchors and dorsal bar.
Urocleidoides macrosoma n. sp. (Figure 5)
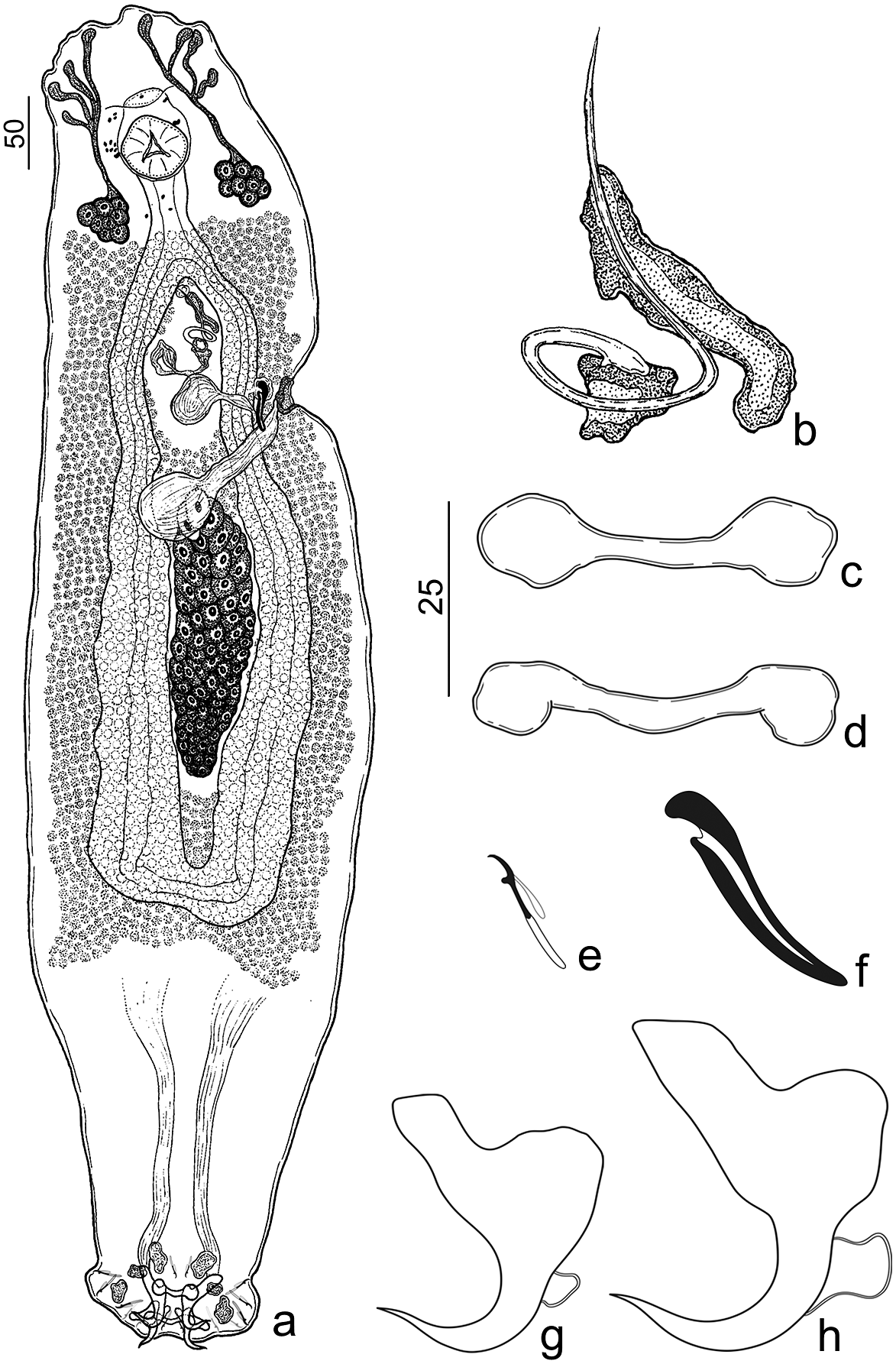
Figure 5. Urocleidoides macrosoma n. sp. a. Holotype, whole body (ventral); b. copulatory complex; c. ventral bar; d. dorsal bar; e. hooks; f. vaginal sclerite; g. dorsal anchor; h. ventral anchor. Scales: a. 50μm scale, b–h. 25μm scale.
Type host. Hoplias malabaricus (Bloch) (Characiformes: Erythrinidae).
Type locality. Vila Segredo – Segredo River (Quatipuru River Basin), Tauari, municipality of Capanema, Pará, Brazil (1°5’32.44”S, 47°5’37.02”W).
Site of infestation. Gills.
Prevalence. 66% (2 of 3 hosts examined).
Average intensity. 1.5 parasites per host.
Specimens deposited. Holotype (CHIOC No. 40204a);2 paratypes (CHIOC No. 40204b–c), 1 hologenophore (CHIOC No. 40204d) .
Molecular sequence data. The partial 28S rDNA (731 bp) and COI mtDNA (733 bp) sequences obtained from one specimen (GenBank accession numbers OR270735 and OR270815, respectively).
Etymology. The specific name of the species derives from the Greek (macro = large + soma = body) and refers to the size of the parasite’s body.
Number of ZooBank. 8A13B412-E0B8-41B3-BE31-F9ED7F409EB5.
Description. (Based on 3 adult specimens – 1 mounted on Gomori’s Trichrome, 1 mounted on Hoyer’s medium, 1 mounted on Gray & Wess). Body elongated, fusiform, robust, total length excluding haptor 795 (737–827; n=3), total width at level of germarium 274 (215–347; n=3) (Figure 5a). Cephalic lobes (4) poorly developed, 2 terminal, 2 bilateral; 4 pairs of head organs; cephalic glands unicellular, posterolateral to pharynx (Figure 5a). Eyes absent; accessory chromatic granules distributed in cephalic region and esophagus (Figure 5a). Pharynx subspherical, muscular, 58 (55–60; n=3) long, 58 (56–62; n=3) wide; esophagus elongated (Figure 5a). Genital pore midventral, anterior to copulatory complex. Genital atrium non-sclerotized. Gonads apparently overlapping. Oviduct, Mehlis’ glands, uterus, egg, testis, prostatic reservoir not observed. Copulatory complex comprising MCO, accessory piece. MCO sclerotized, tubular with one counterclockwise ring, 94 (82–103; n=3) long, base with sclerotized cap, proximal portion of MCO slightly expanded, distal aperture acute (Figure 5b). Accessory piece located in distal portion of MCO, not articulated with base of MCO, comprising an elongated sheath with a groove, which serves as a guide for MCO (Figure 5b). Seminal vesicle with dilated proximal portion, with descending loop followed by ascending loop, distal portion tapered connecting base of MCO. Vaginal pore sinistral, ventro-marginal; vaginal vestibule heavily sclerotized, cup-shaped; vaginal canal muscular. (Figure 5a). Vaginal sclerite 40 (36–46; n=3) long, with longitudinal superficial groove, thumb short, point rounded (Figure 5f). Germarium elongated, fusiform 168 (n=1) long, 55 (n=1) wide. Seminal receptacle subspherical 43 (n=1) long, 46 (n=1) wide. Vitellaria dense, extending from esophagus to confluence of intestinal cecum. Haptor trapezoidal 65 (57–77; n=3) long, 145 (118–162; n=3) wide (Figure 5a). Anchors similar in morphology, robust with elongate superficial root, slightly depressed tip; short deep root, rounded; short shaft; wavy point, extending past level of tip of superficial. Ventral anchor base 31 (29–36; n=3) long, inner 38 (30–42; n=3) long, outer 42 (38–45; n=3) long (Figure 5h). Dorsal anchor base 25 (24–27; n=3) long, inner 34 (32–36; n=3) long, outer 30 (24–37; n=3) long (Figure 5g). Bars similar in morphology, dumbbell-shaped. Ventral bar 38 (36–39; n =3) long (Figure 5c). Dorsal bar 39 (37–39; n=3) long (Figure 5d). Hook pairs similar in morphology with shank divided into two subunits, proximal dilation comprising ½ of shank length, thumb erect, slightly curved shaft, delicate short point, filament hook loop extended to near beginning of shank dilation. Hook pairs 1 and 5 reduced in size, 13 (12–14; n=2) long; pairs 2, 3, 4, 6, 7 17 (17–18; n=2) long (Figure 5e).
Remarks: Urocleidoides macrosoma n. sp. resembles Urocleidoides aimarai Moreira, Scholz & Luque, Reference Moreira, Scholz and Luque2015 by the general morphology of the copulatory complex and anchors. However, they differ from one another mainly due to the morphology of the anchor’s point, bars, germarium, and vaginal sclerite. In U. macrosoma n. sp., anchors have a wavy point, and both bars are dumbbell-shaped, whereas U. aimarai has anchors with evenly curved shaft, point, and a V-shaped ventral bar and rod-shaped dorsal bar with a smooth anteromedial projection. In addition, Urocleidoides macrossoma n. sp. exhibits an elongated and fusiform germarium (bacilliform germarium in U. aimarai) and a vaginal sclerite with a short and rounded point (tapered distal portion of the vaginal sclerite and robust rod in U. aimarai).
Urocleidoides nataliapasternakae n. sp. (Figure 6)

Figure 6. Urocleidoides nataliapasternakae n. sp. a. Holotype, whole body (ventral); b. copulatory complex; c. vaginal sclerite; d. vagina; e. dorsal bar; f. ventral bar; g. hook pair 6; h. hook pair 1; i. hook pair 5; j. hook pairs 2, 3, 4, and 7; k. dorsal anchor; l. ventral anchor. Scales: a. 25μm scale, b–l. 25μm scale.
Type host. Brachyhypopomus brevirostris (Steindachner) (Gymnotiformes: Hypopomidae).
Type locality. Balneário Aracu (Guamá River Basin), municipality of Ourém, Pará, Brazil (1°34’1.02”S, 47° 9’52.35”W).
Site of infestation. Gills.
Prevalence. 50% (2 of 4 hosts examined).
Average intensity. 6.5 parasites per host.
Specimens deposited. Holotype (CHIOC No. 40206a); 9 paratypes (CHIOC No. 40206b–f, 40207a–d), 1 hologenophore (CHIOC No. 40206g) .
Representative DNA sequence. The partial 28S rDNA (766 bp) and COI mtDNA (764 bp) sequences obtained from one specimen (GenBank accession numbers OR270733 and OR270823, respectively).
Etymology. The specific name of the species is a tribute to the biologist and writer Natalia Pasternak Taschner in recognition and admiration for her valuable work of scientific dissemination and communication.
Number of ZooBank. BF8B6560-747D-43CC-B6E6-08FADCE6FA86.
Description. (Based on 10 adult specimens – 4 mounted on Gomori’s Trichrome, 6 mounted on Hoyer’s medium). Body elongated, fusiform, total length excluding haptor 145 (97-201; n=8), total width at level of germarium 56 (44–71; n=9) (Figure 6a). Cephalic lobes (4) moderately developed, 2 terminal, 2 bilateral; 3 pairs of head organs; cephalic glands not observed (Figure 6a). Eyes, accessory chromatic granules absent. Pharynx oval, muscular 16 (14–17; n=4) long, 13 (10–15; n=4) wide; esophagus short (Figure 6a). Genital pore midventral, anterior to copulatory complex. Genital atrium non-sclerotized. Gonads apparently overlapping. Oviduct, Mehlis’ glands, uterus, seminal receptacle, seminal vesicle, testis, prostatic reservoir not observed. Copulatory complex comprising MCO, accessory piece. MCO sclerotized, tubular with two and a half counterclockwise rings,102 (92–119; n=5) long, base with sclerotized cap, proximal portion of MCO slightly expanded, distal aperture acute (Figure 6b). Accessory piece located in distal portion of MCO serving as a guide to MCO, not articulated with base of MCO, comprising a dumbbell-shaped elongated sheath (Figure 6b). Vaginal pore medial, ventral; vaginal vestibule heavily sclerotized; vaginal canal sclerotized, convoluted (Figure 6d). Vaginal sclerite with longitudinal superficial groove, thumb erect, point elongated, slightly straight 28 (20–34; n=6) long (Figure 6c). Germarium elongated, fusiform 29 (27–20; n=3) long, 12 (11–12; n=3) wide. Vitellaria dense, extending from esophagus to confluence of intestinal cecum. Haptor hexagonal 44 (30–54; n=9) long, 68 (53–86; n=9) wide (Figure 6a). Ventral anchor with well-developed superficial root; short deep root, rounded; slightly curved long shaft; curved point extending past level of tip of superficial root, base 11 (10–12; n=7) long, inner 32 (29–38; n=7) long, outer 30 (28–34; n=7) long (Figure 6l). Dorsal anchor with long superficial root, slightly depressed at distal surface; short deep root, rounded; shaft short, curved; point with ornaments as shredded sclerotized filaments, base 12 (10–13; n=7) long, inner 22 (20–23; n=6) long, outer 17 (15–18; n=6) long (Figure 6k). Ventral bar 46 (37–52; n=8) long, straight, rod-shaped with enlarged ends (Figure 6f); dorsal bar 41 (31–49; n=8) long, U-shaped with rounded ends (Figure 6e). Hook pair 1 15 (13–16; n=6) long (Figure 6h), shank with proximal dilation comprising ½ of length of shaft, filament hook loop not observed; hook pairs 2, 3, 4, 7 32 (22–37; n=13) long (Figure 6j) with proximal dilation at shank comprising approximately 1/3 of shank length; hook pair 6 55 (47–62; n=6) long (Figure 6g) larger than others, shank with proximal dilation comprising approximately 1/5 of shank length, filament hook loop not observed; hook pair 5 14 (13–15; n=6) long (Figure 6i) reduced, shank without proximal dilation, filament hook loop not observed.
Remarks. Urocleidoides nataliapasternakae n. sp. resembles Urocleidoides carapus from Gymnotus carapo (Gymnotidae: Gymnotiformes), Urocleidoides cultellus from Brachyhypopomus occidentalis (Gymnotiformes: Hypopomidae), and Urocleidoides ramentacuminatus Oliveira et al., 2019 from Schizodon fasciatus (Characiformes: Anostomidae) by possessing ornamentations at the point of the dorsal anchor. However, U. nataliapasternakae n. sp. is easily distinguished from these species by the combination of the following features: an MCO with two and a half counterclockwise rings (five rings in U. cultellus and one ring MCO in U. ramentacuminatus), a U-shaped dorsal bar (slightly recurved dorsal bar with enlarged ends in U. cultellus), a vagina with a medial aperture (sinistro-marginal vaginal pore in U. ramentacuminatus and Urocleidoides carapus), a vaginal canal convoluted (sigmoid in Urocleidoides carapus and straight in U. ramentacuminatus), and the presence of a vaginal sclerite (absence in U. carapus).
Discussion
Since the amendment of the diagnosis of Urocleidoides proposed by Kritsky et al. (Reference Kritsky, Thatcher and Boeger1986), the presence of vaginal sclerite has been used to define the species of the genus. However, the presence or absence of the vaginal sclerite used to validate species of Urocleidoides is still questioned (Mendoza-Franco & Reina, Reference Mendoza-Franco and Reina2008). According to Mendoza-Franco & Reina (Reference Mendoza-Franco and Reina2008), the species of Urocleidoides incertae sedis (i. e., U. advenai Mendoza-Franco & Reina, Reference Mendoza-Franco and Reina2008, U. carapus, U. gymnotus, and U. hypopomi Suriano, Reference Suriano1997) and sensu stricto (i.e., U. cultellus and U. visiofortatus) described from gymnotiform hosts share some morphological characteristics (i. e., absence of eyes in U. advenai, U. carapus, U. gymnotus, U. cultellus, and U. visiofortatus; ornamentations at the point of the dorsal in U. carapus and U. cultellus; vaginal aperture in the midventral position in U. cultellus, U. gymnotus, U. visiofortatus, and U. hypopomi) suggesting that they may be evolutionarily related. However, these authors commented that the main limitation in diagnosing species of Urocleidoides is the absence of phylogenetic analysis. Kmentová et al. (Reference Kmentová, Cruz-Laufer, Pariselle, Smeets, Artois and Vanhove2022) commented that contradictions in the diagnoses of genera and the morphology of monogenoid species can occur even if the species of a genus exhibit the characteristics listed in the most recent diagnoses. Furthermore, morphological similarities between more distantly related lineages of monogenoids can lead to the erection of several so-called ‘catch-all’ genera. Thus, the molecular data approach for phylogenetic reconstruction in monogenoid studies has been used to highlight and solve such taxonomic problems.
In the present study, the taxonomic status of some species of Urocleidoides with and without vaginal sclerite was evaluated through morphological and molecular data. The phylogenetic analyses based on molecular data support the conclusion that some Urocleidoides species considered incertae sedis (sensu Kritsky et al. Reference Kritsky, Thatcher and Boeger1986) are closely related to their sensu stricto congeners (sensu Kritsky et al. Reference Kritsky, Thatcher and Boeger1986) (Figures 1 and 2). For example, the clade formed by species of Urocleidoides parasitizing gymnotiform fishes (U. cultellus, U. uncinus, U. nataliapasternakae n. sp., all sensu stricto species; U. carapus, U. gymnotus, both incertae sedis species) is well supported by both analyses (ML and BI) using partial sequences of the 28S rDNA gene (BI, P = 1 and ML, B = 100) (Figure 1) and the COI mtDNA gene (BI, P = 0.95) (Figure 2). We also found that U. vanini n. sp. (without vaginal sclerite) and U. atilaiamarinoi n. sp. (with vaginal sclerite) parasites of Characiformes (Erythrinidae) are closely related and supported by the 28S rDNA gene analyses (BI, P = 1 and ML, B = 99) (Figure 1).
Zago et al. (Reference Zago, Yamada, Yamada, Franceschini, Bongiovani and da Silva2020) proposed a phylogenetic hypothesis based on molecular data (COI mtDNA) for species of Urocleidoides reported from Characiformes and Gymnotiformes, whose results showed that Urocleidoides strombicirrus (Price & Bussing, 1967) (incertae sedis) from Characiformes hosts is the sister group of the sensu stricto species, U. cultellus and U. uncinus reported for Gymnotiformes fish. Our results with the inclusion of additional taxa corroborate the findings of Zago et al. (Reference Zago, Yamada, Yamada, Franceschini, Bongiovani and da Silva2020), which also supports the hypothesis proposed by Mendoza-Franco and Reina (Reference Mendoza-Franco and Reina2008) by showing that species with and without vaginal sclerite are closely related. Therefore, although the presence of vaginal sclerite is an important diagnostic characteristic for Urocleidoides, it cannot be considered a main characteristic for the species of the genus. Boeger and Vianna (Reference Boeger, Vianna and Thatcher2006) commented that the presence of a vaginal sclerite in Urocleidoides spp. may be associated with its reproductive system. However, the absence of sclerite in some species may indicate an evolutionary modification in the reproductive mode that may have arisen independently or have been lost secondarily within the group.
Secondary loss of morphological structures has already been reported in some groups of monogenoids. For example, Domingues and Boeger (Reference Domingues and Boeger2008), reviewing species of the family Diplectanidae observed that some genera (i. e., Rhabdosynochus Mizelle & Blatz, Reference Mizelle and Blatz1941, Rhamnocercus Monaco, Wood & Mizelle, Reference Monaco, Wood and Mizelle1954, and Rhamnocercoides Luque & Iannacone, Reference Luque and Iannacone1991) do not present the accessory adhesive organ, which is considered an important feature of the family. Through morphological phylogenetic analysis, they concluded that the absence of such a structure in species of these genera might have been lost secondarily and that this loss probably occurred several times within the evolutionary history of Diplectanidae (see Domingues & Boeger Reference Domingues and Boeger2008). Therefore, based on our results using partial sequences of the 28S rDNA and the COI mtDNA genes, we can conclude that U. carapus, U. gymnotus, and U. vanini n. sp., even if devoid of vaginal sclerite, are valid species for the genus.
Oliveira et al. (Reference Oliveira, da Silva, Vieira and Acosta2021) in their phylogenetic analysis based on 28S rDNA have shown that some species of Urocleidoides appear nested with species of Cacatuocotyle Boeger, Domingues & Kritsky, Reference Boeger and Kritsky1997. The results of our 28S rDNA phylogenetic analysis reveal that the clade formed by Urocleidoides spp. has significant support from both analyses (BI and ML). However, an internal clade with low support shows Cacatuocotyle papilionis as a sister group of the species of Urocleidoides that parasitize gymnotiform and erythrinid (Characiformes) fishes (U. brasiliensis, U. naris, and U. macrosoma n. sp.) (Figure 1). Given this context, our results corroborate those of Oliveira et al. (Reference Oliveira, da Silva, Vieira and Acosta2021) and provide evidence that Urocleidoides may represent a non-monophyletic group.
We detected two clades formed by species of Urocleidoides (Figure 1). For species that parasitize Characiformes, we observed four groups related to host families (i. e., Anostomidae, Erythrinidae, and Parodontidae). In contrast, for species that parasitize Gymnotiformes, we found three groups related to the reported species of Hypopomidae and Gymnotidae (Figure 1). The recovery of these clades might be associated with coevolutionary processes that occurred in isolation within each host family. Boeger and Kritsky (Reference Boeger and Kritsky1997) and Desdevises et al. (Reference Desdevises, Morand, Jousson and Legendre2002) observed that, within coevolutionary scenarios, co-speciation events seem to restrict monogenoid lineages to their hosts and that these events occur at higher taxonomic levels (i. e., family or genus), suggesting that broad historical constraints drive close relationships between monogenoids and their hosts.
Finally, the species of Urocleidoides reported from erythrinid fish in the present study did not represent a monophyletic group (28S rDNA) (Figure 1). We found a clade comprising the species reported from H. malabaricus (U. naris, U. brasiliensis, and U. macrosoma n. sp.) and another with the species reported from E. erythrinus and H. unitaeniatus (U. vanini n. sp. and U. atilaiamarinoi n. sp., respectively). These two clades showed a significant genetic divergence, ranging from 19 to 21.5% (192–214 bp). The species U. atilaiamarinoi n. sp. and U. vanini n. sp. are closely related, reflecting proximity with their hosts (see Oliveira et al. Reference Oliveira, Avelino, Abe, Mariguela, Benine, Ortí, Vari and Castro2011). The 28S rDNA tree shows that the species of Urocleidoides that parasitize H. malabaricus are more closely related to the species that occur in gymnotiform fish. However, the relationships between the species of these clades are not supported, as we found low support values in the BI and ML analyses (Figure 1). Therefore, the separation of clades from species that occur in erythrinid fish may be associated with host exchange events, which may be related to diversification events. In addition, the overlapping geographical distribution of their hosts may also contribute to shaping the sharing of these parasites (Braga et al. Reference Braga, Razzolini and Boeger2015). We suggest that the relationships between the species groups of Urocleidoides can be better elucidated in future studies, with the possible inclusion of all species (i. e., sensu stricto and incertae sedis) in the analyses.
Conclusion
The present study contributes to an understanding of the diversity of species in Urocleidoides, expanding the genus to 52 valid species with the description of four new parasitic species of characiform and gymnotiform fishes from South America. Furthermore, the molecular data from partial sequences of the 28S rDNA and COI mtDNA genes used to reconstruct the phylogenetic relationships between species of Urocleidoides permit a better understanding of the relationships between the sensu stricto species and those considered incertae sedis. Moreover, the absence of vaginal sclerite in some species of Urocleidoides can be explained by secondary loss events, which may have occurred several times within the evolutionary history of the group. We also suggest that the presence of the vaginal sclerite alone is insufficient for diagnosing the species of the genus. Finally, future studies may clarify the correct taxonomic status of the other species still considered incertae sedis and thus generate more robust data for a better understanding of the evolutionary history of this host–parasite system.
Acknowledgements
The authors thank Geusivam Soares (UNICAMP) for reviewing early versions of this manuscript and the members of the Laboratório de Sistemática e Coevolução (UFPA) and J-M.F.L. Cunha for assistance during the collecting trips. We thank the anonymous reviewers for their contributions to this manuscript.
Financial support
The present study was partly supported by research grants from the Conselho Nacional de Desenvolvimento Científico e Tecnológico, CNPq – Brazil (CNPq - PROTAX 001/2015-440526/2015-9) and Fundação Amazônia de Amparo a Estudos e Pesquisas, FAPESPA – Brazil (ICAAF 017-2018) to M.V. Domingues. J.F. Santos-Neto was supported by a doctoral scholarship from the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior, CAPES – Brazil (grant# 88882.460123/2019-01). M.V. Domingues received research productivity grants from the Conselho Nacional de Desenvolvimento Científico e Tecnológico (grant #309896/2019-3). The preparation of this manuscript was supported by Programa de Apoio à Publicação Qualificada (PAPQ 2022) of Pró-Reitoria de Pesquisa e Pós-Graduação (PROPESP) of the Universidade Federal do Pará (UFPA).
Competing interest
None.
Ethical standard
All applicable institutional, national, and international guidelines for the care and use of animals were followed. Specimens were collected under the license for collection of biological material (43381) granted by the Instituto Chico Mendes de Conservação da Biodiversidade (ICMBio).











