



Introduction
Ophiocomid brittle stars are abundantly found in shallow waters and coral reefs of all tropical regions (O'Hara et al., Reference O'Hara, Hugall, Cisternas, Boissin, Bribiesca-Contreras, Sellanes, Paulay and Byrne2019). They have fascinating camouflage skills and negative phototaxis under a range of light intensities that is used as visual protection against predators (Hendler, Reference Hendler1984; Hendler and Byrne, Reference Hendler and Byrne1987; O'Hara et al., Reference O'Hara, Byrne and Cisternas2004, Reference O'Hara, Hugall, Cisternas, Boissin, Bribiesca-Contreras, Sellanes, Paulay and Byrne2019).
The taxonomy of Ophiocoma L. Agassiz, 1836, has been uncertain due to morphological characters that are not easily recognized and understood, and characters that can change during growth, a common fact among ophiuroids (Sumida et al., Reference Sumida, Tyler, Gage and Norrevang1998; Stöhr, Reference Stöhr2005; Stöhr et al., Reference Stöhr, Conand and Boissin2008). The genus is currently considered monophyletic, and the presence of an ornate egg envelope has been regarded as a synapomorphy of the Ophiocoma clade (O'Hara et al., Reference O'Hara, Hugall, Cisternas, Boissin, Bribiesca-Contreras, Sellanes, Paulay and Byrne2019).
Before the review by O'Hara et al. (Reference O'Hara, Hugall, Cisternas, Boissin, Bribiesca-Contreras, Sellanes, Paulay and Byrne2019), Ophiocoma genus was represented by three species in Brazilian waters, O. pumila Lütken, 1856, O. wendtii Müller & Troschel, Reference Müller and Troschel1842 and O. echinata (Lamarck, Reference Lamarck1816) (Tommasi, Reference Tommasi1970; Tommasi and Aron, Reference Tommasi and Aron1988). Such authors in 2019 transferred O. pumila to Ophiocomella A.H. Clark, 1939, O. wendtii to Ophiomastix Müller & Troschel, 1842, and O. echinata was the only remaining Ophiocoma species recorded from the Brazilian coast so far, whose distribution ranges from Florida (~20°N) to Bahia (~12°S) (Tommasi, Reference Tommasi1970; Manso, Reference Manso1993). Localities along the Brazilian coast include the states of Ceará (Albuquerque, Reference Albuquerque1986), Paraíba (Rathbun, Reference Rathbun1879), Pernambuco (Tommasi, Reference Tommasi1970; Lima and Fernandes, Reference Lima and Fernandes2009), Alagoas (Miranda et al., Reference Miranda, Lima, Sovierzoski and Correia2012) and Bahia (Tommasi, Reference Tommasi1970; Manso et al., Reference Manso, Alves and Martins2008).
During a long-term survey of the remote oceanic archipelago of Trindade and Martin Vaz, located off the coast of southeastern Brazil (Tavares et al., Reference Tavares, Carvalho and Mendonça2017), a wealth of specimens of Ophiocoma were obtained. A taxonomic study based on morphological and molecular analyses from both insular and coastal specimens revealed that the Trindade and Martin Vaz samples belong to a new species, whereas the coastal specimens belong to O. echinata. The new species is herein described, illustrated and a redescription of O. echinata is provided (https://zoobank.org/NomenclaturalActs/6943697D-DB27-4904-AB9F-5EF4243E37B).
Materials and methods
Abbreviations: BA, Bahia; CBS, Caribbean Sea; ES, Espírito Santo; RI, Reunion Island; SPSPA, São Pedro and São Paulo Archipelago; USP, University of São Paulo; MDBio, Museum of Biological Diversity, Zoology Area (ZUEC); MZUSP, Zoology Museum of USP.
Morphological observations and comparisons with original descriptions of each one of the genera of the Ophiocomidae family allowed assigning the studied specimens to the Ophiocoma genus, which has alternation in the number of arm spines and enlargement of the upper arm spines (Devaney, Reference Devaney1970) as observed in Ophiocoma sp. TMV (Trindade and Martin Vaz Archipelago). Thereafter, morphological comparisons were made among Ophiocoma sp. TMV and seven other Ophiocoma species, recently reviewed by O'Hara et al. (Reference O'Hara, Hugall, Cisternas, Boissin, Bribiesca-Contreras, Sellanes, Paulay and Byrne2019): Ophiocoma aethiops Lütken, Reference Lütken1859; O. anaglyptica Ely, Reference Ely1944; O. cynthiae Benavides-Serrato & O'Hara, 2008; O. echinata (Lamarck, Reference Lamarck1816); O. erinaceus Müller & Troschel, Reference Müller and Troschel1842; O. schoenleinii Müller & Troschel, Reference Müller and Troschel1842; and O. scolopendrina (Lamarck, Reference Lamarck1816), with the aim of verifying if Ophiocoma sp. TMV corresponds to any of the seven described species of Ophiocoma. These comparisons were made based on the original descriptions and most recent publications (Say, Reference Say1825; Müller and Troschel, Reference Müller and Troschel1842; Matsumoto, Reference Matsumoto1917; Ely, Reference Ely1944; Devaney, Reference Devaney1970; Tommasi, Reference Tommasi1970; O'Hara et al., Reference O'Hara, Stöhr, Hugall, Thuy and Martynov2018, Reference O'Hara, Hugall, Cisternas, Boissin, Bribiesca-Contreras, Sellanes, Paulay and Byrne2019).
Study site and data collection
The studied specimens were collected by the project ProTrindade/CNPq – TMV and belong to the scientific collections of the ZUEC and MZUSP. However, additional samples were collected in the Northeastern Brazil (NB), specifically in Salvador – BA.
The TMV is located in the southwestern Atlantic, approximately 1.200 km east of Vitoria, ES. The Trindade Island (20°30′S, 29°20′W) is formed by a single large island of 13.5 km2, while Martin Vaz Archipelago (20°30′S, 28°51′W) is located 47 km east of Trindade Island and formed by several smaller islands (Figure 1) (Marques et al., Reference Marques, Ulbrich, Ruberti and Tassinari1999; Almeida, Reference Almeida2007; Anker et al., Reference Anker, Tavares and Mendonça2016; Alitto et al., Reference Alitto, Amaral, Oliveira, Serrano, Seger, Guilherme, Di Domenico, Christensen, Lourenço, Tavares and Borges2019). The expeditions executed by the ProTrindade/CNPq project occurred between the years 2012 and 2019, and the material was collected by scuba diving at depths of 0.5–30 m, which resulted in 397 lots of Ophiuroidea.
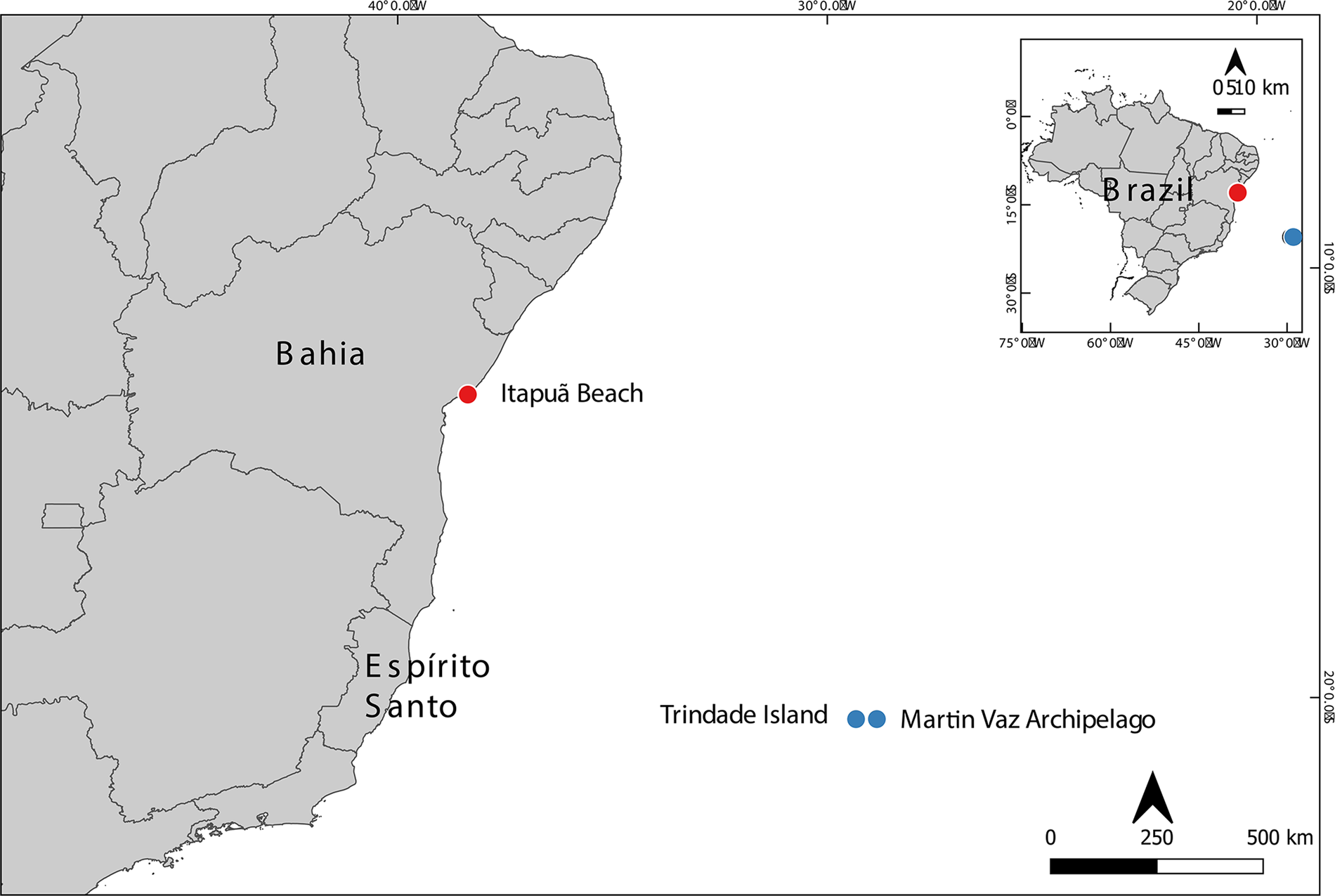
Figure 1. Study areas: Trindade Island and Martin Vaz Archipelago, Espírito Santo and Itapuã Beach, Bahia.
Salvador is located in the state of Bahia, on the northeast coast of Brazil. The Itapuã beach (12°57′S, 38°21′W), where the specimens were obtained, consists of a wide area of exposed reefs, which receive little energy from the shore (Alves and Cerqueira, Reference Alves and Cerqueira2000). All the collections were done manually between the rock fragments at a depth of approximately 10 cm.
Some additional specimens were examined and were important for our delimitation: Saint Thomas Danish West Indies, US Virgin Islands, type locality of Ophiocoma echinata, sampled in 1963 (Voucher ZUEC OPH 1953) and Carrie Bow Cay Belize (Voucher ZUEC OPH 1893), sampled in 1989. Both vouchers contain one specimen and are in 70% ethanol.
Delimitation of the morphotypes
In order to delimit the morphotypes studied, the specimens were initially separated based on the traditionally used external morphological characters, such as the type of disc cover, number, size and characteristics of spines, and tentacle scales, among others. Intraspecific differences of each delimited group were also analysed.
Morphological characters
External morphology
The arm spines were counted from the first to the 21st segment on both sides of each arm (Benavides-Serrato and O'Hara, Reference Benavides-Serrato and O'Hara2008) and the count of the granules was done in three frames measuring 0.2 mm2, in the central area of the disc, from photos taken using the program AxioVision VS. 40.4.8.20 (Carl Zeiss Microscopy, Germany) attached to a stereomicroscope ZEISS Discovery. The average of these three frames was then multiplied by five, creating an estimate of the number of disc granules in an area of 1 mm2 (Stöhr et al., Reference Stöhr, Boissin and Hoareau2013). Classical morphological taxonomy was adopted for external and internal morphological studies with the following specialized literature (Borges et al., Reference Borges, Monteiro and Amaral2002; Borges and Amaral, Reference Borges, Amaral, Amaral, Rizzo and Arruda2005; Stöhr et al., Reference Stöhr, O'Hara and Thuy2012, Reference Stöhr, Boissin and Hoareau2013; Thuy and Stöhr, Reference Thuy and Stöhr2016; Hendler, Reference Hendler2018; O'Hara et al., Reference O'Hara, Stöhr, Hugall, Thuy and Martynov2018).
Morphology of the ossicles
In order to describe and analyse intra- and interspecific morphological differences and similarities, we studied the microstructures of the arm ossicles, such as dorsal, ventral and lateral arm plates, as well as vertebrae. These were taken from a fragment of the arm (between the fifth and tenth segment) of an adult from both coastal and insular specimens of each morphotype. For other ossicles, such as oral, dental and genital plates, the entire disc or part thereof was used. Each arm or disc fragment was immersed in regular household bleach (NaClO) until the tissues were removed (Stöhr et al., Reference Stöhr, Conand and Boissin2008). The ossicles were then washed in distilled water and ethanol, air-dried and arranged on aluminium stubs, sputtered with gold for observation under a scanning electron microscope (SEM) model JEOL JSM5800LV at the Electron Microscopy Laboratory of the Institute of Biology at the University of Campinas (UNICAMP).
The terminology used in the description of the ossicles essentially followed Thuy and Stöhr (Reference Thuy and Stöhr2011, Reference Thuy and Stöhr2016), Alitto et al. (Reference Alitto, Bueno, Guilherme, Di Domenico, Christensen and Borges2018), O'Hara et al. (Reference O'Hara, Stöhr, Hugall, Thuy and Martynov2018) and Hendler (Reference Hendler2018). Detailed images of ossicles descriptions are shown in Figure 2.
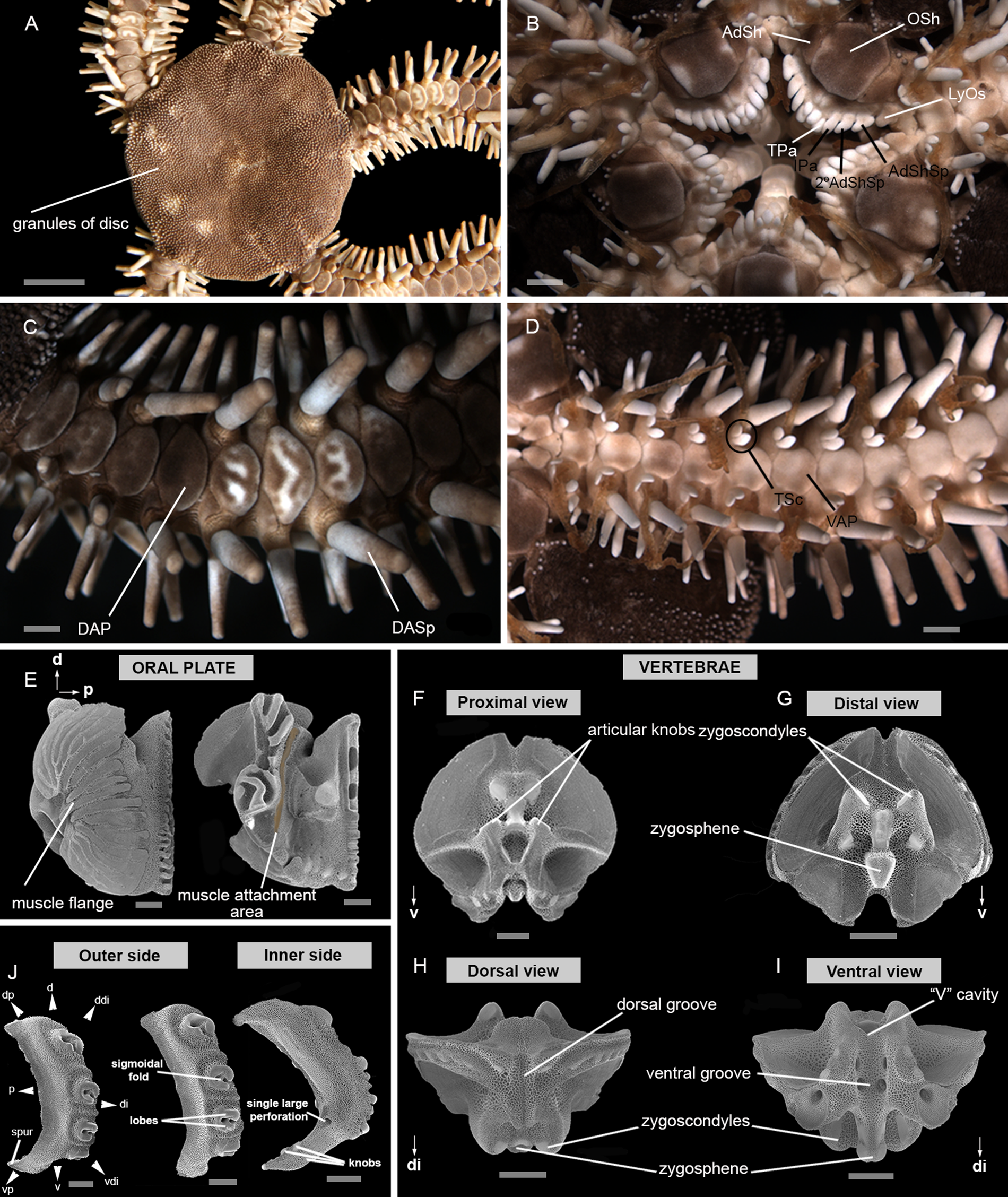
Figure 2. Main characters used in the description: (A) dorsal view; (B) ventral view; (C) detail of dorsal arm; (D) detail of ventral arm; (E) oral plate – abradial and adradial; (F) vertebrae ossicle – proximal surface; (G) vertebrae ossicle – distal surface; (H) vertebrae ossicle – dorsal surface; (I) vertebrae ossicle – ventral surface; (J) lateral arm plate – outer and inner side. 2° AdShSp, secondary adoral shield spine; AdSh, adoral shield; AdShSp, adoral shield spine; d, dorsal; DAP, dorsal arm plate; DASp, dorsalmost arm spine; ddi, dorso-distal; di, distal; dp, dorso-proximal; IPa, infradental papilla; LyOs, Lyman's ossicle; Osh, oral shield; p, proximal; TPa, tooth papilla; TSc, tentacle scale; v, ventral; VAP, ventral arm plate; vdi, ventro-distal; vp, ventro-proximal. Scale bar: stereomicroscope photographs A, 5 mm; B–D, 1 mm; SEM images E–J, 500 μm.
Morphometry
Morphometry and linear discriminant analysis (LDA) were used to refine and improve the previously established morphological studies of external characters and distinguish groups. Measurements were taken using the program AxioVision VS. 40.4.8.20 (Carl Zeiss Microscopy) attached to a stereomicroscope ZEISS Discovery V20 for specimens and structures smaller than 10 mm and/or with a digital calliper for those larger than 10 mm. A detailed list of the measured morphological characters is shown in Table 1 and Figure 3.
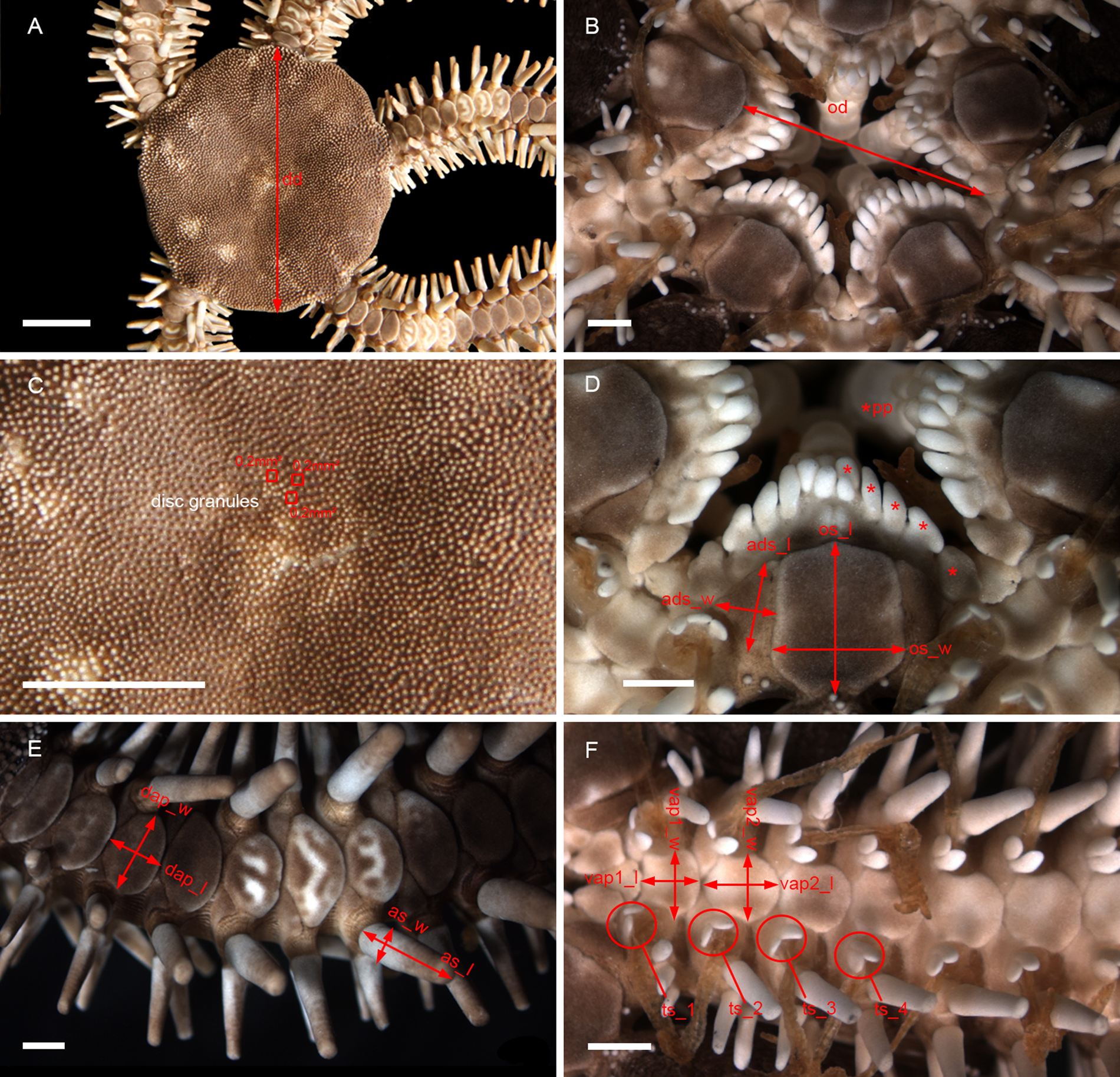
Figure 3. Main characters used in morphometry: (A) dorsal view; (B) detail of the oral view; (C) disc granules; (D) detail of the jaw; (E) detail of dorsal arm; (F) detail of ventral arm. dd, disc diameter; as_l, length of arm spine; as_w, width of arm spine; od, oral diameter; os_l, length of oral shield; os_w, width of oral shield; ads_l, length of adoral shield; ads_w, width of adoral shield; dap_l, length of dorsal arm plate; dap_w, width of dorsal arm plate; vap1_l, length of first ventral arm plate; vap1_w, width of first ventral arm plate; vap2_l, length of second ventral arm plate; vap2_w, width of second ventral arm plate; pp, oral papillae; ts_1, tentacle scale of first arm segment; ts_2, tentacle scale of second arm segment; ts_3, tentacle scale of third arm segment; ts_4, tentacle scale of fourth arm segment. Scale bar: stereomicroscope photographs A and C, 5 mm; B, D, E and F, 1 mm.
Table 1. Abbreviations and definitions of morphological characters measured for morphometric analysis

LDA was used to determine the differences between the groups in the classification (Venables and Ripley, Reference Venables and Ripley2002). A correlation matrix was constructed and the variables that were significantly correlated were removed with a threshold value of 0.9, aiming to avoid multicollinearity among morphological characters. Additionally, the variables near zero variances were removed (Kuhn and Johnson, Reference Kuhn and Johnson2013). All analyses were conducted in R version 4.0.4 (2021-02-15) using software packages, such as MASS (Venables and Ripley, Reference Venables and Ripley2002; R Development Core Team, 2016), ggord (Beck, Reference Beck2022), vegan (Oksanen et al., Reference Oksanen, Blanchet, Friendly, Kindt, Legendre, McGlinn, Minchin, O'Hara, Simpson, Solymos, Stevens, Szoecs and Wagner2019), tidyverse (Wickham et al., Reference Wickham, Averick, Bryan, Chang, McGowan, François, Grolemund, Hayes, Henry, Hester, Kuhn, Pedersen, Miller, Bache, Müller, Ooms, Robinson, Seidel, Spinu, Takahashi, Vaughan, Wilke, Woo and Yutani2019) and cowplot (Wilke, Reference Wilke2019).
A total of 79 specimens of Ophiocoma were measured, 63 of TMV and 16 of NB. In order to perform the LDA analysis, a total of 20 morphological characters were used, and the analysis was shown to be effective in discriminating between the two species of Ophiocoma.
Molecular characters
DNA extraction, amplification and sequencing
To analyse DNA sequences and compare them with the other sequences obtained from different species of Ophiocoma, the total genomic DNA was extracted according to the protocol previously established and tested by Alitto et al. (Reference Alitto, Amaral, Oliveira, Serrano, Seger, Guilherme, Di Domenico, Christensen, Lourenço, Tavares and Borges2019), where tube feet from the arm and/or gonads were soaked in two changes of Tris-EDTA (TE) buffer for 30 min. The sample was then macerated in 50 μl of TE buffer with a pestle. Another 450 μl of TE buffer and 300 μl of Chelex 100 10% (Walsh et al., Reference Walsh, Metzger and Higuchi1991) were added, and then the samples were incubated for 60 min at 55°C, boiled in a water bath for 8 min and, after reaching room temperature, vortexed and centrifuged for 1 min. The supernatant (300 μl) containing the genomic DNA was removed and incubated at −20°C.
Fragments of the mitochondrial gene 16S were amplified by polymerase chain reaction (PCR) through the primers 16S Sofi F and Sofi R (Pérez-Portela et al., Reference Pérez-Portela, Almada and Turon2013). PCR reactions were performed with a final volume of 25 μl, with 2.5 μl of reaction buffer (Taq Buffer), 2.5 μl of dNTP, 4 μl of MgCl2 (1.5 mM, Invitrogen), 1 μl of each of the primers and 1 μl of Taq DNA polymerase, provided by the companies ‘Exxtend Soluções em Oligos’, Paulínia-SP, Brazil and ‘LCR’, Cosmópolis-SP, Brazil. The amount of DNA and water was variable, aiming to complete the total volume (25 μl).
The thermal cycling consisted of a single step at 94°C for 7 min, which was followed by 39 cycles (denaturation at 94°C for 60 s, annealing at 50–56°C for 70 s and extension at 72°C for 80 s) and a final extension at 72°C for 5 min on a thermal cycler (Eppendorf Mastercycler®). For purification, the Promega Kit was used, and subsequently, the samples were sent to the sequencing service of Myleus Biotechnology. All the sequences are deposited in GenBank (OM388509 to OM388511).
Phylogenetic analyses
For the phylogenetic analysis of Ophiocoma, the 16S fragment of the species Ophiocomella ophiactoides (H.L. Clark, 1900), also from the Ophiocomidae family, was used as the outgroup (all samples used from GenBank are shown in Table 2).
Table 2. GenBank access number for DNA sequences used for phylogenetic analyses

NMRJ, National Museum of Rio de Janeiro, Brazil; URUN, University of Reunion Island, Reunion.
Two approaches were used for phylogenetic analyses: maximum parsimony (MP) and Bayesian inference (BI). The nodes in the cladogram obtained by the MP were tested by the bootstrap sampling test (Felsenstein, Reference Felsenstein1985), based on 1000 pseudoreplicates using the new technology (Alitto et al., Reference Alitto, Amaral, Oliveira, Serrano, Seger, Guilherme, Di Domenico, Christensen, Lourenço, Tavares and Borges2019).
Two simultaneous analyses were performed, with four chains in each one (three heated and one cold). A total of 3,000,000 generations were executed in each run, with one tree sampled every 100 generations. A consensus topology with the subsequent probability for each node was produced after the disposal of the first trees generated (initial 25%). The value of average standard deviation of split frequencies was less than 0.01. Phylogenetic trees were visualized and edited using the FIGTREE v.1.4.3 program (http://tree.bio.ed.ac.uk/software/figtree/).
The genetic distances between and within the different groups were estimated by p-distance using the program MEGA v. X (Kumar et al., Reference Kumar, Stecher, Li, Knyaz and Tamura2018).
Results
Here, we present the results of the multiple comparative analyses, including data from external and microstructural morphology, and morphometry, in order to understand whether the two morphotypes of the genus Ophiocoma (Ophiocoma sp. TMV and Ophiocoma sp. NB) found in Brazil do or do not belong to the same species and whether they belong to one (or several) of the known species. A total of 106 specimens were analysed from two regions: (i) TMV (90 specimens) and (ii) NB (16 specimens). They were separated into two morphotypes: Ophiocoma sp. TMV and Ophiocoma sp. NB.
Morphometry
A correlation was found between the number of tentacle scales from the first to the fourth arm segment (ts_1, ts_2, ts_3 and ts_4), and for that reason, this character was excluded from the analysis. Only one axis (LD1) was necessary to separate the two morphotypes (Figure 4), and the characters with the highest positive coefficients were the width of the second ventral arm plate (vap2_w) and the length of the first ventral arm plate (vap1_l), and those with the lowest negative coefficients were the length of the oral shield (os_l) and the oral diameter (od).

Figure 4. Linear discriminant analysis (LDA) analysis based on 20 morphological variables (described in Table 1) from two morphotypes: Ophiocoma sp. TMV and Ophiocoma sp. NB. (A) Histogram of the LDA scores; (B) density plot of the values of the discriminant function for the samples from different morphotypes. dd, disc diameter; disc_granule, granules of disc; as_l, length of arm spine; as_w, width of arm spine; as, arm spine of first segment; as2, arm spine of second segment; as3, arm spine of third segment; as10, arm spine of tenth segment; od, oral diameter; os_l, length of oral shield; os_w, width of oral shield; ads_l, length of adoral shield; ads_w, width of adoral shield; dap_l, length of dorsal arm plate; dap_w, width of dorsal arm plate; vap1_l, length of first ventral arm plate; vap1_w, width of first ventral arm plate; vap2_l, length of second ventral arm plate; vap2_w, width of second ventral arm plate; pp, oral papillae.
Morphometrically, Ophiocoma sp. NB differs from Ophiocoma sp. TMV in having greater width of the second ventral arm plate (Figure 5A), length of the first ventral arm plate (Figure 5B), oral diameter (Figure 5C) and length of the oral shield (Figure 5D) than Ophiocoma sp. TMV.

Figure 5. Boxplots generated by the LDA analysis of two morphotypes: Ophiocoma sp. TMV and Ophiocoma sp. NB. Boxplot of positive morphological variables: (A) vap2_w and (B) vap1_l; and negative (C) od and (D) os_l. od, oral diameter; os_l, length of the oral shield; vap1_l, length of first ventral arm plate; vap2_w, width of second ventral arm plate.
Molecular characters
A sequence of 361 bp from three individuals was obtained for 16S, two from TMV and one from NB. The phylogenetic reconstruction, obtained by BI, shows two clades, A and B (Figure 6), a similar result to the analysis with MP.

Figure 6. Phylogenetic reconstruction of the 16S gene obtained with Bayesian inference. The numbers above the branches represent posterior probabilities. For clades also inferred in the MP analysis, bootstrap values (%) are provided (below the branches).
Clade A (in orange), which contains O. echinata from the Caribbean Sea and Ophiocoma sp. NB, presented a high support value for both the BI and the MP (0.99 and 99%, respectively). Clade B (in blue), which contains specimens of Ophiocoma sp. from the TMV, also showed a high support value for the two analyses, BI and MP (1 and 99%, respectively).
In BI, the standard deviation of the split frequencies between the two runs reached a value lower than 0.005 at two million generations. The analysis of four executions resulted in similar likelihood scores, with ESS (Effective Sample Size)>200, as verified using TRACER. The most suitable model for molecular evolution was SYM + G, chosen based on the Akaike information criterion and hierarchical likelihood ratio tests.
The genetic distances within clade A (TMV) were low (0.77%), while the genetic distance between clades A and B was 12.9–13.1%. These results corroborate the morphological and morphometric analysis in the separation of these two morphotypes. The genetic distances of the 16S sequences are shown in Table 3.
Table 3. Genetic distance (%) between and within groups of 16S

#, value of the genetic distance within the group not calculated because only one sequence was considered in the analysis.
Species delimitation
All analyses indicated the separation of the two morphotypes, Ophiocoma sp. TMV and Ophiocoma sp. NB. Divergences were found between external morphology (number of tentacles scales), microstructural characters (dorsal and ventral arm plates, dental plates and especially dorsalmost arm spine), morphometry and molecular data (16S and genetic distances). These results corroborate the initial hypothesis of the existence of two species of Ophiocoma at the Brazilian coast, Ophiocoma trindadensis sp. nov. and O. echinata. In the following, we present a taxonomic key for both species, a detailed description of the new species, and a redescription of O. echinata.
Taxonomic key
Two oval tentacle scales on each pore of the first arm segment, then one on all following segments. Three to four arm spines. Dorsal arm spine almost two times as long as wide, with lateral edges tapered. Dental plate long, with dorsal and ventral portion of the same width. Dorsal arm plates with lateral edges tapered. Ventral arm plates with distal portion wider than proximal portion and distal edge straight…………O. trindadensis sp. nov.
Two oval tentacle scales on each pore, except in the final third of the arm. Three to five arm spines. Dorsal arm spine robust, almost four times as long as wide. Dental plate long with dorsal portion wider than ventral portion. Dorsal arm plates with lateral edges rounded. Ventral arm plates with distal portion as wide as the proximal one and distal edge rounded…………O. echinata.
Systematics
Order OPHIACANTHIDA O'Hara et al., Reference O'Hara, Hugall, Thuy, Stöhr and Martynov2017
Family OPHIOCOMIDAE Ljungman, Reference Ljungman1867
Genus Ophiocoma L. Agassiz, Reference Agassiz1836
Ophiocoma trindadensis Serrano, Damiano, Alitto and Borges sp. nov.
(Figures 7 and 8)
https://zoobank.org/135d9966-bf8a-4b77-851a-41b5aa779b3f
https://zoobank.org/NomenclaturalActs/6943697D-DB27-4904-AB9F-5EF4243E37B
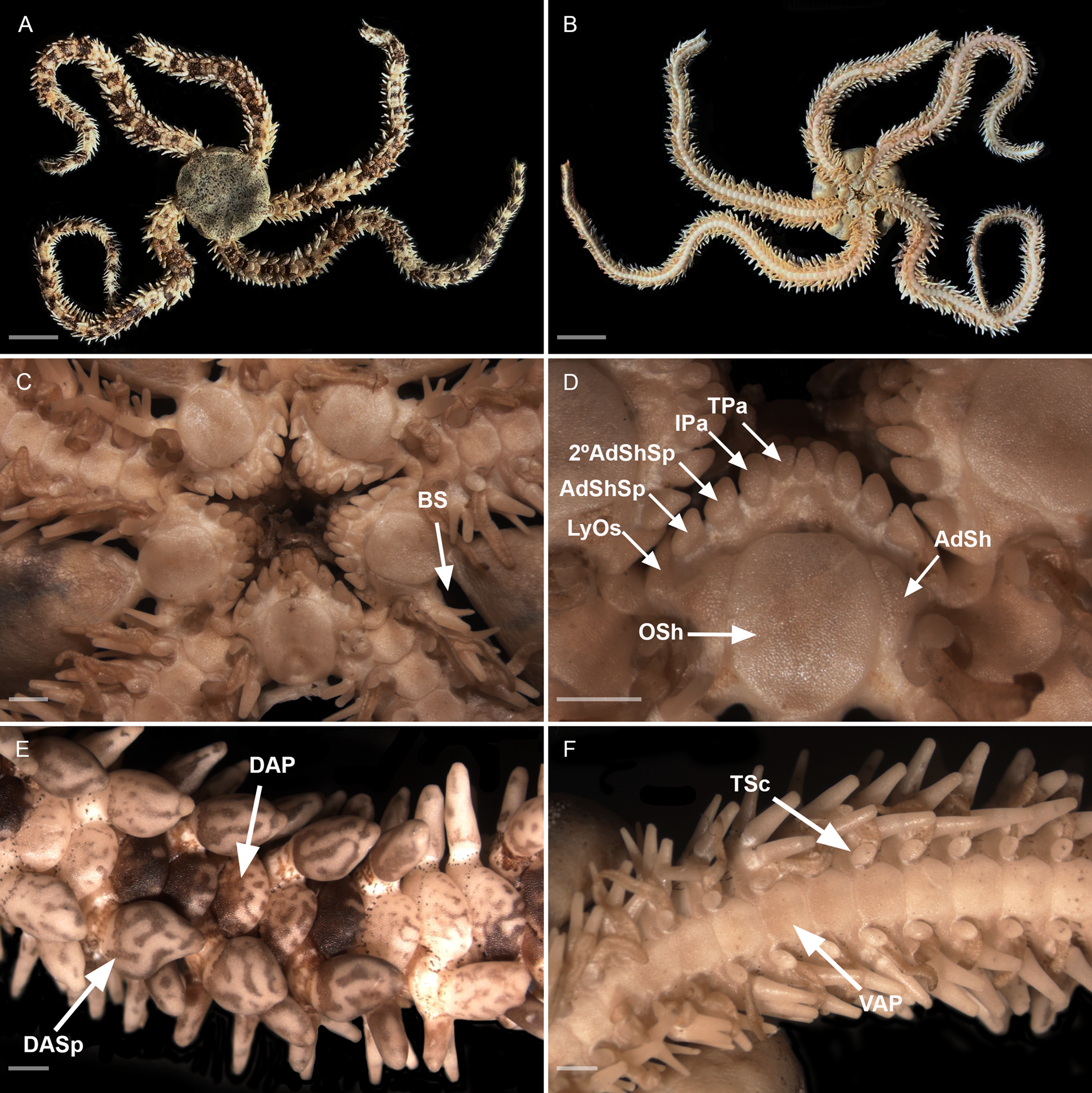
Figure 7. Ophiocoma trindadensis sp. nov. Sample ZUEC OPH 3095 (19.3 mm dd). (A) Dorsal view; (B) ventral view; (C) detail of the oral view; (D) detail of the jaw; (E) detail of dorsal arm; (F) detail of ventral arm. 2° AdShSp, secondary adoral shield spine; AdSh, adoral shield; AdShSp, adoral shield spine; BS, bursal slits; DAP, dorsal arm plate; DASp, dorsalmost arm spine; IPa, infradental papilla; LyOs, Lyman's ossicle; Osh, oral shield; TPa, tooth papilla; TSc, tentacle scale; VAP, ventral arm plate. Scale bar: A and B, 10 mm; C–F, 1 mm.
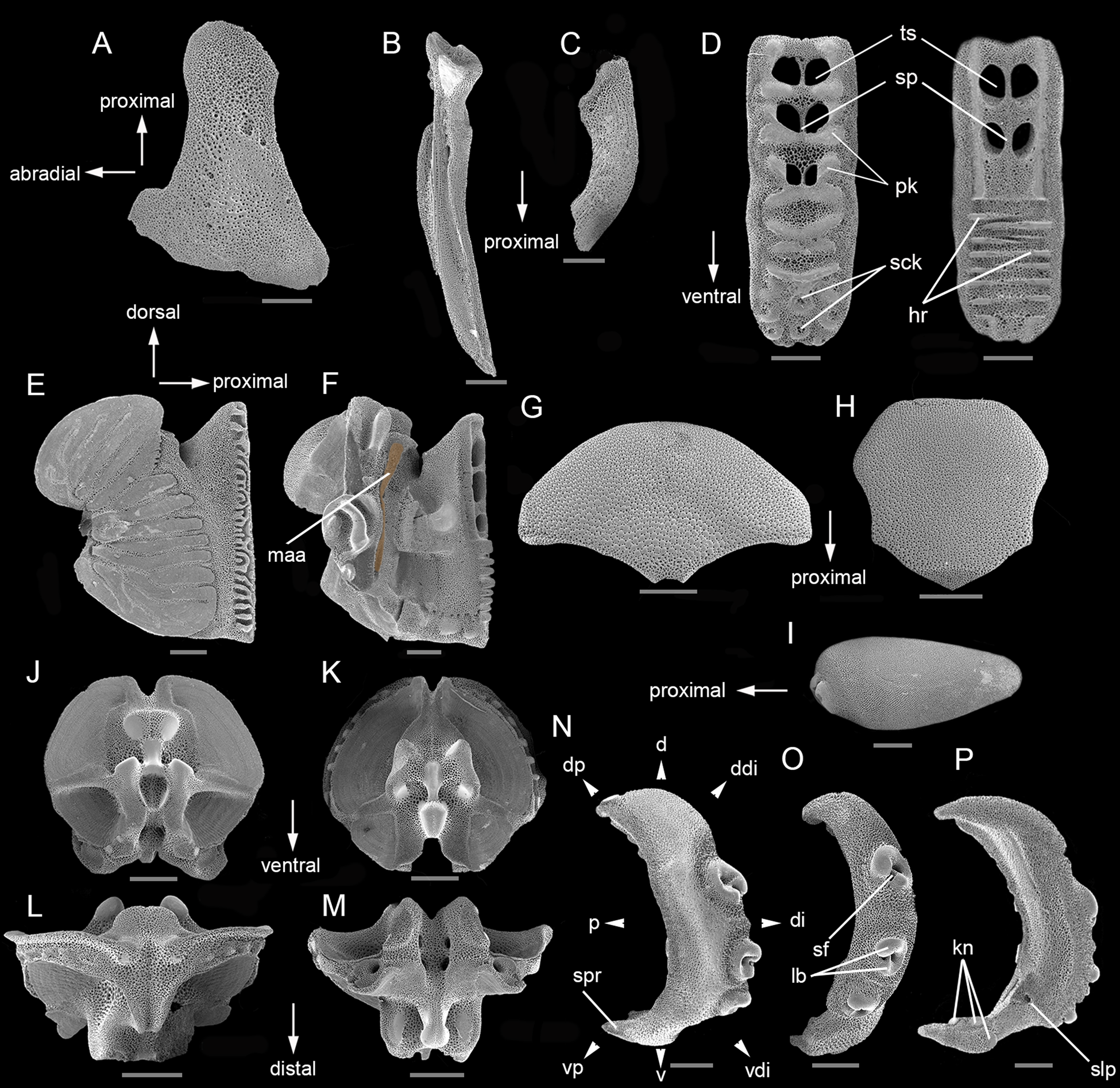
Figure 8. Microstructural characters of Ophiocoma trindadensis sp. nov. Sample ZUEC OPH 3095 (19.3 mm dd): (A) radial shield – outer side; (B) adradial genital plate; (C) abradial genital plate; (D) dental plate – outer and inner side; (E) oral plate – adradial; (F) oral plate – abradial (highlighted muscle attachment area); (G) dorsal arm plate – outer side; (H) ventral arm plate – outer side; (I) dorsal arm spine; (J) vertebra – proximal surface; (K) vertebra – distal surface; (L) vertebra – dorsal surface; (M) vertebra – ventral surface; (O) lateral arm plate – spine articulations. Sample MZUSP 2768 (19 mm dd): (N) lateral arm plate – outer side; (P) lateral arm plate – inner side. d, dorsal; ddi, dorso-distal; di, distal; dp, dorso-proximal; hr, horizontal ridge; kn, knob; lb, lobes; maa, muscle attachment area; p, proximal; pk, protruding knobs; sck, sockets; sf, sigmoidal fold; slp, single large perforation; sp, septum; spr, spur; ts, tooth sockets; v, ventral; vdi, ventro-distal; vp, ventro-proximal. Scale bars: A–P, 500 μm.
Type locality. Trindade and Martin Vaz Archipelago, ES, BRAZIL.
Size range. dd: 1.7–19.3 mm.
Material examined. 90 specimens (dd: 1.7–19.4 mm). See specimens information (Supplementary Table 1).
Holotype: MZUSP 1973, adult, 19.4 mm dd, collected from Trindade Island, Ponta da Calheta, Espírito Santo, Brazil, 20°30′18.72″S 29°18′31.67″W at a depth of 17.7 m; coll. Joel B. Mendonça, 3 August 2015.
Paratypes: Trindade Island, ES, Brazil; coll. Joel B. Mendonça: MZUSP 1916 (1 spm; 12.1 mm dd); MZUSP 2745 (1 spm; 11.3 mm dd); MZUSP 2746 (1 spm; 5.1 mm dd); MZUSP 2747 (1 spm; 10.9 mm dd); MZUSP 2757 (1 spm; 8.7 mm dd); MZUSP 2809 (1 spm; 3.7 mm dd); MZUSP 2772 (1 spm; 8 mm dd); MZUSP 2810 (1 spm; 3.9 mm dd); MZUSP 2774 (1 spm; 5.1 mm dd); MZUSP 2775 (1 spm; 2.9 mm dd); MZUSP 2776 (1 spm; 3.1 mm dd); MZUSP 2777 (1 spm; 11 mm dd); MZUSP 2780 (1 spm; 9.3 mm dd); ZUEC OPH 3066 (1 spm; 8.5 mm dd); ZUEC OPH 3067 (1 spm; 9.7 mm dd); ZUEC OPH 3068 (1 spm; 9.1 mm dd); ZUEC OPH 3069 (1 spm; 12.6 mm dd); ZUEC OPH 3070 (1 spm). Martin Vaz Archipelago, Martin Vaz Island, ES, Brazil, coll. Joel B. Mendonça: MZUSP 3101 (1 spm; 10 mm dd). See specimens information.
Additional specimens: see specimens information (Supplementary Table 1).
Diagnosis
Disc and radial shields completely covered by sparse and rounded granules. Dorsalmost arm spines as wide as long or almost as wide as long, rounded and robust. Dorsal arm plates contiguous, overlapping proximally. Two oval tentacle scales on each pore of the first arm segment, then one on all following segments.
Description
Disc (dd: 19.3 mm). Circular, completely covered dorsally by sparse and rounded granules, extending ventrally. About 13 granules per mm2 (Figure 7A).
Mouth. Oral shields rectangular with rounded edges, 1.2 times as long as wide. Adoral shield triangular, twice as wide as long, separated proximally (Figure 7C, D). At each half-jaw a sequence of five papillae: Lyman's ossicle large, partially covered by a triangular adoral shield spine, 1.3 times as long as wide; secondary adoral shield spine a rounded scalene triangle-shaped, 1.2 times as long as wide; infradental papilla oval, 1.7 times as long as wide; at ventralmost portion of dental plate, tooth papilla oval and shorter than the other papillae (Figure 7D).
Arms. Dorsal arm plates contiguous, overlapping proximally. Ventral arm plates hexagonal. First ventral arm plate with the proximal edge covered by Lyman's ossicle (Figure 7C). Second ventral arm plate, 1.5 times as wide as long, distally concave with an angular lateral edge on both sides. Lateral edge of other ventral arm plates with concavity and a convex/rounded distal edge (Figure 7F). The number of arm spines on each side from the first arm segment until 21st, are arranged in the following sequence: 
Three to four arm spines. The number of spines alternates at the same arm segment (on one side with three, on the other side with four). Between third and 21st arm segments, the dorsalmost arm spines are more robust and rounded (Figure 7E), as wide as long or almost as wide as long (with 2.2 mm of length and 1.5 mm of width) in segments with four spines. Two oval tentacle scales on each pore of the first arm segment and only one on the rest (Figure 7F).
Morphological variations. Granules: the larger specimens (19.2 and 19.3 mm dd) can present 4–13 granules per mm2 in the centre of the disc, while the smallest specimens (1.7–4.8 mm dd) can present 61–113 granules per mm2. Arm spines: some specimens with dorsalmost spines more rounded and bigger than others, and ventralmost spines bifid and flattened. Tentacle scales: varying number on the same individual and between specimens, one and two tentacle scales on the first arm segment in approximately 35%, and only two tentacle scales in 65% of individuals. Approximately 5% have two tentacle scales on the second arm segment.
Microstructure of ossicles (Figure 8)
Material examined. Sample ZUEC OPH 3095 (dd: 19.3 mm).
Radial shield. Abradial distal edge incised (Figure 8A).
Genital plates. Adradial plate: long, thin and with a condyle and groove (Figure 8B). Abradial plate: longer than half the length of the adradial plate, sickle-shape (Figure 8C).
Dental plate. General outline: long, with dorsal and ventral portion of the same width. Outer side: dorsalmost portion with two tooth sockets divided by a thin septum into two halves, and in the central portion one tooth socket divided into two small holes. Holes of the tooth sockets longer than wide, surrounded by protruding knobs; tooth papillae sockets on ventralmost portion. Inner side: dorsalmost portion with two tooth sockets divided by a thin septum into two halves, longer than wide. A series of horizontal ridges from the median to the ventralmost portion (Figure 8D).
Oral plate. General outline: 1.2 times as long as wide. Abradial: presents a large and well-defined muscle flange with horizontal and diagonal striations (Figure 8E). Adradial: muscle attachment area arranged vertically, long and tapering, spoon-shaped (Figure 8F).
Dorsal arm plates. Fan-shaped, 1.8 times as wide as long; lateral edges tapered (Figure 8G).
Ventral arm plates. Heptagonal, as wide as long, distal portion 1.25 times as wide as long as the proximal one; distal edge straight, proximal angle with straight edges and lateral edges concave for the tentacle openings (Figure 8H).
Dorsal arm spine. Almost two times as long as wide, with lateral edges tapered (Figure 8I).
Vertebrae. Zygospondylous articulation: Proximal side with two articular knobs and a dense stereom, positioned in the middle of the dorsal portion (Figure 8J). Distal side with zygosphene fused with a pair of parallel zygocondyles (Figure 8K). Dorsal side with a groove not projecting beyond the zygocondyles (Figure 8L). Ventral side with a strong depression in the groove upper proximal half, a ‘V’ cavity in the proximal edge and zygosphene slightly projecting beyond the ventral edge of zygocondyles (Figure 8M).
Lateral arm plates. General outline: Arched position (wrapped around the arm); ventral portion projecting ventro-proximalwards (Figure 8N). Outer surface ornamentation: trabecular intersections protruding to form knobs larger than stereom pores on most of the outer surface (Figure 8N). Outer proximal edge: surface lined discernible band of different stereom structure, over most of the proximal edge; present an oblique, elongated spur on ventro-proximal tip of outer surface (Figure 8N). Arm spine articulations: three, on elevated portion not bordered proximally by ridge; directly adjacent to the distal edge; arranged over entire distal lateral arm plate; dorsalwards increasing in size; distance between spine articulations dorsalwards increasing. Lobes merged at their proximal tips by smooth connection; one lobe clearly larger than the other, not parallel, shifted, bent, with tilted orientation; stereom with perforations; sigmoidal fold fully developed (Figure 8O). Inner side: dominated by more or less continuous ridge; three separate knobs, with differentiated stereom, on the ventral tip; single large perforation (Figure 8P).
Morphological variations. Dorsal arm plate: some dorsal arm plate can be irregular, with one edge longer than the other. Dental plate: can present two or three tooth sockets divided by a septum into two halves. Lateral arm plate: some lateral arm plate can present four spine articulations.
Etymology
The species name ‘trindadensis’ alludes to the location where the species was collected.
Ecology and distribution
Ophiocoma trindadensis sp. nov. was collected from rubble bottom. It has also been sampled in corals and associated with calcareous algae. The present study samples occurred at depths ranging from 8 to 23.5 m, only at the Trindade and Martin Vaz Archipelago.
Ophiocoma echinata (Lamarck, Reference Lamarck1816)
(Figures 9 and 10)
Type locality. Antilles
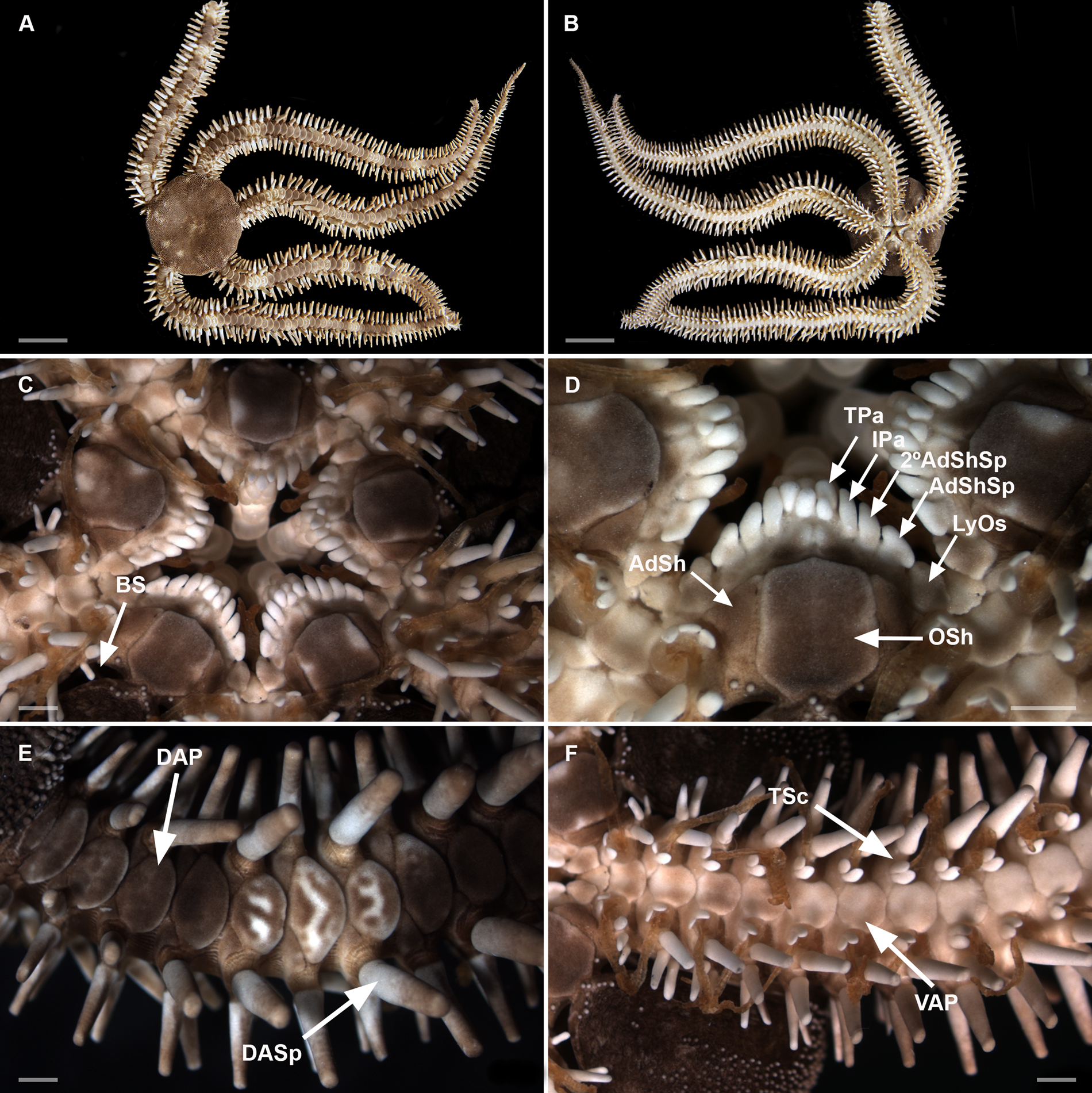
Figure 9. Ophiocoma echinata. Sample ZUEC OPH 1538 (19.3 mm dd). (A) Dorsal view; (B) ventral view; (C) detail of the oral view; (D) detail of the jaw; (E) detail of dorsal arm; (F) detail of ventral arm. 2° AdShSp, secondary adoral shield spine; AdSh, adoral shield; AdShSp, adoral shield spine; BS, bursal slits; DAP, dorsal arm plate; DASp, dorsalmost arm spine; IPa, infradental papilla; LyOs, Lyman's ossicle; Osh, oral shield; TPa, tooth papilla; TSc, tentacle scale; VAP, ventral arm plate. Scale bars: A and B, 10 mm; C–F, 1 mm.
Size range. dd: 4.9–22.7 mm.
Material examined. 16 specimens (dd: 4.9–22.7 mm). Salvador, Bahia, Brazil: ZUEC OPH 1538 (1 spm; 19.3 mm dd); ZUEC OPH 3071 (1 spm; 22.6 mm dd); ZUEC OPH 3072 (1 spm; 19.5 mm dd); ZUEC OPH 3073 (1 spm; 22.7 mm dd); ZUEC OPH 3074 (1 spm; 22.5 mm dd); ZUEC OPH 3075 (1 spm; 17.4 mm dd); ZUEC OPH 3076 (1 spm; 17.3 mm dd); ZUEC OPH 3077 (1 spm; 8.2 mm dd); ZUEC OPH 3078 (1 spm; 15.2 mm dd); ZUEC OPH 3079 (1 spm; 4.9 mm dd); ZUEC OPH 3080 (1 spm; 13.6 mm dd); ZUEC OPH 3081 (1 spm; 15.8 mm dd); ZUEC OPH 3082 (1 spm; 15.2 mm dd); ZUEC OPH 3083 (1 spm; 13.7 mm dd); ZUEC OPH 3084 (1 spm; 14.3 mm dd); ZUEC OPH 3085 (1 spm; 9.2 mm dd). See specimens information (Supplementary Table 1).
Diagnosis
Disc and radial shields sparsely covered by rounded granules. Dorsalmost arm spines rounded, longer than wide. Dorsal arm plates not contiguous. Two oval tentacle scales on each pore, except in the final third of the arm.
Description
Disc (dd: 19.3 mm). Circular, sparsely covered dorsally by rounded granules, extending ventrally. About 15 granules per mm2 (Figure 9A).
Mouth. Oral shields rectangular with angular margins, 1.2 times as long as wide. Adoral shield triangular, 1.8 times as wide as long, separated proximally (Figure 9C, D). At each half-jaw a sequence of five papillae: Lyman's ossicle large, 1.5–1.8 times as long as wide; adoral shield spine triangular 1.4 times as long as wide; secondary adoral shield spine elongated, 2.2 times as long as wide; infradental papilla elongated, 1.6 times as long as wide ventralmost portion of dental plate, an oval tooth papilla, shorter than the other papillae (Figure 9D).
Arms. Dorsal arm plates not contiguous. Ventral arm plates hexagonal. First ventral arm plate with the proximal edge covered by Lyman's ossicle (Figure 9C). Second ventral arm plate, almost as long as wide, distally concave with an angular lateral edge on both sides. Lateral edge of other ventral arm plates with concavity and a convex/rounded distal edge (Figure 9F). The number of arm spines on each side, from the first arm segment until 21st, are arranged in the following sequence: 
For counting the number of dorsal arm spines, another specimen of similar size (19.5 mm dd) was used, which had the most preserved dorsal arm spines and only four entire arms. Three to five cylindrical arm spines. The number of spines alternates in the same arm segment (on one side with three, on the other side with four). Between fifth and 14th arm segments the dorsalmost arm spines are more robust (Figure 9E), longer than wide (3.4 mm in length and 1.5 mm in width) in segments with four spines. Two and three oval tentacle scales on each pore, except in the final third of the arm (Figure 9F).
Morphological variations. Granules: the larger specimens (22.6 and 22.7 mm dd) can present 10–13 granules per mm2 in the centre of the disc, while the smallest specimens (4.9 and 8.2 mm dd) can present 27–38 granules per mm2. Tentacle scales: two oval tentacle scales on the first arm segment in 88% of specimens, three in 6% and four tentacle scales on the first arm segment in approximately 6%.
Microstructurure of ossicles (Figure 10)
Material examined. Sample ZUEC OPH 1538 (dd: 19.3 mm)
Radial shield. Abradial distal edge concave, distal edge more than twice as wide as proximal end (Figure 10A).

Figure 10. Microstructural characters of Ophiocoma echinata. Sample ZUEC OPH 1538 (19.3 mm dd): (A) radial shield – outer side; (B) genital plate – adradial; (C) genital plate – abradial; (D) dental plate – outer and inner side; (E) oral plate – adradial; (F) oral plate – abradial (highlighted muscle attachment area); (G) dorsal arm plate – outer side; (H) ventral arm plate – outer side; (I) dorsal arm spine; (J) vertebra– proximal surface; (K) vertebra – distal surface; (L) vertebra – dorsal surface; (M) vertebra – ventral surface. Sample ZUEC OPH 3072 (19.5 mm dd): (N) lateral arm plate – outer side; (O) lateral arm plate – spine articulation; (P) lateral arm plate – inner side. d, dorsal; ddi, dorso-distal; di, distal; dp, dorso-proximal; hr, horizontal ridge; kn, knob; lb, lobes; maa, muscle attachment area; p, proximal; pk, protruding knobs; sck, sockets; sf, sigmoidal fold; slp, single large perforation; sp, septum; spr, spur; ts, tooth sockets; v, ventral; vdi, ventro-distal; vp, ventro-proximal. Scale bar: A–E and I–P, 500 μm; F and G, 200 μm.
Genital plates. Adradial plate: long, thin and with a condyle and groove (Figure 10B). Abradial plate: longer than half the length of the adradial plate; sickle-shape (Figure 10C).
Dental plate. General outline: long with dorsal portion wider than ventral portion. Outer side: dorsalmost portion with two tooth sockets divided by a thick septum into two halves, and in the central portion one tooth socket divided into two small holes. Holes of the tooth sockets longer than wide, surrounded by protruding knobs; tooth papillae sockets on ventralmost portion. Inner side: dorsalmost portion with two tooth sockets divided by a thick septum into two halves longer than wide. A series of horizontal ridges from the median to the ventralmost portion (Figure 10D).
Oral plate. General outline: 1.3 times as long as wide. Abradial: presents a large and well-defined muscle flange with horizontal and diagonal striations (Figure 10E). Adradial: muscle attachment area arranged vertically, long and tapering, spoon-shaped (Figure 10F).
Dorsal arm plates. Fan-shaped, 1.4 times as wide as long; lateral edges rounded (Figure 10G).
Ventral arm plates. Heptagonal, as wide as long, distal portion as wide as the proximal one; distal edge rounded, proximal angle with straight edges and lateral edges concaves for the tentacle openings (Figure 10H).
Dorsal arm spine. Robust, almost four times as long as wide (Figure 10I).
Vertebrae. Zygospondylous articulation: Proximal side with two articular knobs and dense stereom, positioned in middle of dorsal portion (Figure 10J). Distal side with zygosphene fused with a pair of parallel zygocondyles (Figure 10K). Dorsal side with a groove not projecting beyond the zygocondyles (Figure 10L). Ventral side with a strong depression in the groove upper proximal half, a ‘V’ cavity in the proximal edge and zygosphene projecting beyond ventral edge of zygocondyles (Figure 10M).
Lateral arm plates. General outline: Arched position (wrapped around the arm); ventral portion projecting ventro-proximalwards (Figure 10N). Outer surface ornamentation: trabecular intersections protruding to form knobs larger than stereom pores on most of the outer surface (Figure 10N). Outer proximal edge: surface lined discernible band of different stereom structure, over most of the proximal edge; present an oblique, elongated spur on ventro-proximal tip of outer surface (Figure 10N). Arm spine articulations: four, on elevated portion not bordered proximally by ridge; directly adjacent to the distal edge; arranged over entire distal edge; dorsalwards increasing in size; distance between spine articulations dorsalwards increasing. Lobes merged at their proximal tips by smooth connection; one lobe clearly larger than the other, not parallel, shifted, bent, with tilted orientation; stereom with perforations; sigmoidal fold fully developed (Figure 10O). Inner side: dominated by more or less continuous ridge; three separate knobs, with differentiated stereom, on the ventral tip; single large perforation (Figure 10P).
Morphological variations. Dorsal arm plate: some dorsal arm plates have lateral edges more elongated than rounded. Dental plate: with two or three tooth sockets divided by a septum. Lateral arm plate: some lateral arm plates can present three spine articulations.
Ecology and distribution
The present study samples were collected at Itapuã beach, from exposed reefs at depths of 10 cm. Ophiocoma echinata from other localities occurred at depths from 0.1 to 49 m, occurring on substrates such as coral and sandstone reefs. In Brazil: Bahia (Tommasi, Reference Tommasi1970; Manso et al., Reference Manso, Alves and Martins2008), Ceará (Albuquerque, Reference Albuquerque1986), Pernambuco (Tommasi, Reference Tommasi1970; Lima and Fernandes, Reference Lima and Fernandes2009), Alagoas (Miranda et al., Reference Miranda, Lima, Sovierzoski and Correia2012) and Paraíba (Rathbun, Reference Rathbun1879). Atlantic Ocean: From Florida to Brazil (Tommasi, Reference Tommasi1970).
Discussion
As a starting point, two morphotypes of Ophiocoma were characterized from initial morphological analyses, occurring in Brazil: Ophiocoma sp. NB and Ophiocoma sp. TMV. To investigate whether the two morphotypes corresponded to one or two species, an integrative analysis of morphological (external morphology, morphometry and microstructural characters) and molecular characters (16S) was performed. All results suggest that the analysed specimens belong to two distinct species: O. echinata from NB and O. trindadensis sp. nov. from TMV. All the specimens of O. echinata from NB were further compared with O. echinata from the type locality, belonging to the collection of Ophiuroidea of the MDBio.
Ecologically, the species O. echinata and O. trindadensis sp. nov. were found in similar types of substrates and in the same bathymetric distribution. However, the geographic distribution of the species is distinct, since O. echinata is restricted to the continental coastal region, in which it is widely distributed (Ceará to Bahia), while O. trindadensis is exclusive to the remote oceanic archipelago of Trindade and Martin Vaz.
Ophiocoma echinata is the only species of the genus that occurs at the Brazilian coast and the analysed specimens of O. trindadensis sp. nov. do not correspond morphologically to it. However, although many Ophiocoma species share spherical granular cover and thick and evident spines (Lamarck, Reference Lamarck1816), they differ in number of tentacle scales, and size and shape of the dorsalmost arm spines. According to Say (Reference Say1825), O. echinata has two oval and equal tentacle scales along the entire arm, and dilated dorsalmost arm spines, which are almost oval. Unlike O. echinata, O. trindadensis sp. nov. has two tentacle scales in the first arm segment and only one in the others, besides very robust dorsalmost arm spines, which are more rounded than in O. echinata.
As much as the dorsalmost arm spines of Ophiocoma anaglyptica Ely, Reference Ely1944 are described as swollen and enlarged, they are characterized as bottle-shaped, expanded in the middle but slightly compressed, narrowing abruptly to form a short neck (Ely, Reference Ely1944). In addition, this species from the Pacific Ocean has flattened disc granules, and two tentacle scales on all but the first segments, which can support up to three tentacle scales (Ely, Reference Ely1944; Devaney, Reference Devaney1970). Lyman (Reference Lyman1861) described Ophiocoma molaris (synonymized as Ophiocoma scolopendrina), with two tentacle scales in the first arm segment and then one, but dorsalmost arm spines are slender, long and tapering, being three to three and a half times as long as dorsal arm plates. The descriptions of Ophiocoma erinaceus Müller & Troschel, Reference Müller and Troschel1842 and Ophiocoma cynthiae Benavides-Serrato & O'Hara, Reference Benavides-Serrato and O'Hara2008 do not indicate similarities with O. trindadensis sp. nov., since both have two tentacle scales and O. erinaceus also has bright red tubular feet in live specimens (Müller and Troschel, Reference Müller and Troschel1842; Benavides-Serrato and O'Hara, Reference Benavides-Serrato and O'Hara2008).
Although Ophiocoma schoenleinii Müller & Troschel, Reference Müller and Troschel1842 and Ophiocoma aethiops Lütken, Reference Lütken1859 have a single tentacle scale, except in the first segments of the arm (Müller and Troschel, Reference Müller and Troschel1842; Matsumoto, Reference Matsumoto1917; Koehler, Reference Koehler1922; Devaney, Reference Devaney1970), both do not have dilated dorsalmost arm spines as O. trindadensis sp. nov. (Koehler, Reference Koehler1922; Fatemi et al., Reference Fatemi, Jamili, Valinassab and Kuranlu2010). In addition, O. schoenleinii occurs in the Indian and the Pacific Ocean while O. aethiops occurs only in the Pacific Ocean.
The study of the ossicles allowed to observe differences between the morphotypes analysed. The dental plates of O. trindadensis sp. nov. have a thin septum, the ventral arm plates are wider distally than proximally, and the dorsal arm plates have more prominent edges, creating a curvature towards the proximal portion. In contrast, O. echinata from NB differs by presenting more pronounced concavity in the ventral arm plates and dorsal arm plates are more oval.
The LDA analysis also corroborated the separation of specimens into two groups. Ophiocoma echinata from NB, in addition to presenting longer first ventral arm plates, and wider second ventral arm plates, has a longer oral shield and the oral diameter is larger than in O. trindadensis sp. nov.
Regarding molecular analysis, from 16S sequences, differences were also found between the morphotypes. In both phylogenetic inferences, MP and BI, two clades were observed, one for the TMV species and the other for Ophiocoma sp. NB and specimens from the Antilles. The genetic distances between these clades were considered high (12.9–13.1%).
Until the present moment, O. echinata and O. trindadensis sp. nov. are the only two species of Ophiocoma recorded for Brazil, the latter being known only from the Trindade and Martin Vaz Archipelago, so far.
Supplementary material
The supplementary material for this article can be found at https://doi.org/10.1017/S0025315423000383.
Acknowledgements
We are grateful to all professors and researchers involved in this work: especially Luciana Bolsoni and Karin Seger (LABESC), and Gabriela Granadier (EQUILAB), as well as Vanessa Silva Vicente. We also thank MZUSP for specimens donated and the access to MEV equipment, and assistance provided by the Electron Microscope Laboratory (LME/UNICAMP).
Author contributions
Analysing the data: Helena Serrano, Renata A. S. Alitto, Cecília J. S. Damiano, Pablo D. B. Guilherme, Michela Borges. Carrying out the study: Helena Serrano, Renata A. S. Alitto, Cecília J. S. Damiano, Michela Borges. Designing the study: Helena Serrano, Renata A. S. Alitto, Cecília J. S. Damiano, Michela Borges. Formulating the research questions: Helena Serrano, Renata A. S. Alitto, Cecília J. S. Damiano, Michela Borges. Funding acquisition: Joel B. Mendonça, Marcos Tavares, Walter R. P. Cerqueira, Michela Borges. Interpreting the findings: Helena Serrano, Renata A. S. Alitto, Cecília J. S. Damiano, Pablo D. B. Guilherme, Walter R. P. Cerqueira, Michela Borges. Writing the article: Helena Serrano, Renata A. S. Alitto, Cecília J. S. Damiano, Pablo D. B. Guilherme, Michela Borges.
Financial support
This work was supported by the São Paulo Research Foundation (FAPESP, Process no. 2019/01620-9), and CAPES (Coordenação de Aperfeiçoamento de Pessoal de Nível Superior – Brasil) – Finance Code 001. We also thank FAPESP for the scientific initiation scholarship (Process no. 2019/05997-0), post-doctoral scholarship (2020/09031-0) and to the BIOTA-Thematic Projects (Process no. 2018/10313-0 and 2016/05843-4). Furthermore, we thank the financial support of the ProTrindade Project (Process no. 2016/50373-6), CNPq (Conselho Nacional de Desenvolvimento Científico e Tecnológico – Process no. 402186/2016-8 and 443278/2019-9), the Brazilian Navy (1st District) and SECIRIM (Interministerial Secretariat for Marine Resources) for all the support provided in Trindade and Martin Vaz. The material used to refer to the Trindade and Martin Vaz Archipelago was contribution 14 of the ProTrindade Marine Invertebrates Project.
Competing interest
None.
Ethical standards
According to the Ministry of the Environment of Brazil, the activity of accessing the Genetic Heritage was registered in SisGen (Sistema Nacional de Gestão do Patrimônio Genético e do Conhecimento Tradicional Associado), with the purpose of accessing the Genetic Heritage of the species Ophiocoma sp., for the purpose of access in Research. Registration number: A3014E.















