


Introduction
Nogami (Reference Nogami1966, Reference Nogami1967) published the first studies on Cambrian conodonts recovered from North China. An (Reference An1981, Reference An1982) and An et al. (Reference An, Zhang, Xiang, Zhang, Xu, Zhang, Jiang, Yang, Lin, Cui and Yang1983) greatly expanded systematic studies on Cambrian conodonts of northern and northeastern China. Wang (Reference Wang1985) and Chen and Gong (Reference Chen and Gong1986) focused on conodonts across the Cambrian–Ordovician boundary in northern and northeastern China. Wang and Luo (Reference Wang and Luo1984), An and Zheng (Reference An and Zheng1990), and Zhao et al. (Reference Zhao, Zhang and Xiao2000) found only sporadic Cambrian conodonts in northwestern China.
An (Reference An1982) used a more biostratigraphic approach. He made good use of the favorable geological conditions in northern and northeastern China, mainly in four key sections in Liaoning, Hebei, and Shandong. There, he founded conodont zones of the Zhangxia Formation (middle Cambrian) through the Fengshan Formation (upper Cambrian). Paraconodont zones below the Fengshan Formation were the first to be proposed for this interval anywhere in the world.
An et al. (Reference An, Du, Gao, Chen and Li1981) also initiated studies of Cambrian conodonts in South China, although they focused on conodonts across the Cambrian–Ordovician boundary. They found Cordylodus proavus Müller, Reference Müller1959 and other important elements, such as Monocostodus sevierensis (Miller, Reference Miller1969), in the dolostone in the top part of the Sanyoudong Formation in Yichang, Hubei. An et al. (Reference An, Du, Gao, Chen and Li1981) first proposed that the Cambrian–Ordovician boundary should be drawn at ~10 m below the top of the Sanyoudong Formation, which previously had been considered to be entirely upper Cambrian. Dong (Reference Dong1983, Reference Dong1985) restudied conodonts in the interval across the Cambrian–Ordovician boundary in Yichang, where he found Cordylodus intermedius Furnish, Reference Furnish1938 and Cordylodus angulatus Pander, Reference Pander1856. Dong founded three conodont assemblage zones in these Yichang strata and proposed that the Cambrian–Ordovician boundary there should be drawn at the base of the Monocostodus sevierensis–Cordylodus intermedius Assemblage Zone, which is 14 m below the top of the Sanyoudong Formation in Yichang. Dong (Reference Dong1987) recognized the Proconodontus Zone, Cordylodus proavus Zone, and C. intermedius Zone in Chuxian, Anhui, South China. Based on conodonts from western Zhejiang and Chuxian, Anhui in South China, Ding et al. (Reference Ding, Cao, Bao and Yang1993a, Reference Ding, Chen, Zhang, Cao, Bao and Yangb) distinguished the Proconodontus Zone and the C. proavus Zone. Western Hunan has a favorable geological setting for systematic studies of Cambrian conodonts in South China because of relatively continuous carbonate deposition, but conodont research there was in a pioneer state before the late 1980s. Xi-ping Dong began studying Cambrian through lowermost Ordovician conodont biostratigraphy and taxonomy in western Hunan in 1985. An and Hu (Reference An and Hu1985) and An (Reference An1987) studied two sections, namely at Luoyixi in Guzhang County and Likouzui in Fenghuang County. Because they recovered only a few Cambrian conodonts, it was not possible to propose conodont zones. Recently, Bagnoli and colleagues did excellent work on the Cambrian conodonts of South China (Qi et al., Reference Qi, Bagnoli and Wang2006; Bagnoli et al., Reference Bagnoli, Qi and Wang2008; Bagnoli and Qi, Reference Bagnoli and Qi2011).
One difficulty in biostratigraphic study of Cambrian conodonts has been the lack of a detailed zonation based on paraconodonts below the upper upper Cambrian. A regionally applicable zonation based on earliest euconodonts, which first appear in the upper upper Cambrian, was largely established in North America in the 1980s and has been improved subsequently (Miller et al., Reference Miller, Evens, Loch, Ethington, Stitt, Holmer and Popov2003). This zonation based on early euconodonts played an important role in delimiting the Cambrian–Ordovician boundary and should prove to be useful in defining the terminal stage of the Cambrian (Miller, Reference Miller1980, Reference Miller1981, Reference Miller1987, Reference Miller1988; Miller et al., Reference Miller, Taylor, Stitt, Ethington, Hintze and Taylor1982, Reference Miller, Ethington, Evans, Holmer, Loch, Popov, Repetski, Ripperdan and Taylor2006, Reference Miller, Evens, Freeman, Ripperdan and Taylor2011, Reference Miller, Evens, Freeman, Ripperdan and Taylor2014a; Forty et al., Reference Fortey, Landing and Skevington1982; Taylor and Landing, Reference Taylor and Landing1982; Nowlan, Reference Nowlan1985; Bagnoli et al., Reference Bagnoli, Barnes and Stevens1987; Barnes, Reference Barnes1988; Fåhræus and Roy, Reference Fåhræus and Roy1993; Landing, 1979, Reference Landing1993; Landing and Westrop, 2006; Landing et al., Reference Landing, Barnes and Stevens1986, Reference Landing, Westrop and van Aller Hernick2003, Reference Landing, Westrop and Keppie2007, Reference Landing, Westrop and Adrain2011; Ji and Barnes, Reference Ji and Barnes1994; Taylor and Repetski, Reference Taylor and Repetski1995; Nicoll et al., Reference Nicoll, Miller, Nowlan, Repetski and Ethington1999; Parsons and Clark, Reference Parsons and Clark1999; Cooper et al., Reference Cooper, Nowlan and Williams2001; Pyle et al., Reference Pyle, Barnes and Mcanally2007). Since 1982, several of these zones and subzones have been successively recognized in North China (An, Reference An1982; An et al., Reference An, Zhang, Xiang, Zhang, Xu, Zhang, Jiang, Yang, Lin, Cui and Yang1983; Wang, Reference Wang1985; Chen and Gong, Reference Chen and Gong1986; Nowlan and Nicoll, Reference Nowlan and Nicoll1995), South China (Dong, Reference Dong1985, Reference Dong1987; Dong et al., Reference Dong, Repetski and Bergström2004c), Australia (Nicoll, Reference Nicoll1990, Reference Nicoll1991; Nicoll and Shergold, Reference Nicoll and Shergold1991), Kazakhstan (Dubinina, Reference Dubinina2000), Mexico (Landing et al., Reference Landing, Westrop and Keppie2007), New Zealand (Wright and Cooper, Reference Wright and Cooper1994), Turkey (Göncüoğlu and Kozur, Reference Göncüoğlu and Kozur2000), and South Korea (Lee, Reference Lee2001, Reference Lee2002a, Reference Leeb, Reference Lee2004, Reference Lee2008a, Reference Leeb; Lee et al., Reference Lee, Lee, Miller and Jeong2009).
Despite all of this published work, conodonts in the middle Cambrian and most of the upper Cambrian have not been recognized worldwide as useful index fossils. There are several reasons for this. First, trilobites have been, and remain, the key fossils for biostratigraphic subdivision and correlation of Cambrian rocks, and most biostratigraphic studies have been centered on this group. Other groups, including conodonts, were largely neglected until the late 1960s. Second, the routine technique of etching samples with acetic acid for conodonts is suitable for processing only carbonate rocks. Conodonts are generally rare or absent in many dolostones, so only limestones are suitable in most cases for large-scale processing. However, continuously deposited and well-exposed middle Cambrian to Lower Ordovician limestone successions occur only in a few countries, such as China. Third, there has been a tendency among many conodont workers to consider the Cambrian paraconodont taxa to be stratigraphically long ranging and hence of limited use for stratigraphic subdivision and correlation.
In Europe, the well-studied middle to upper Cambrian sections in Scandinavia are stratigraphically very condensed, and the trilobite biostratigraphy has been well studied. These strata have particularly great potential for study of paraconodonts. Conodont faunas in these successions were described in a magnificent monograph by Müller and Hinz (Reference Müller and Hinz1991). However, because the aim of their study was taxonomic rather than biostratigraphic, they focused their sampling on fossiliferous limestone nodules in the black shale succession. The age of each sample was subsequently established based on the trilobite taxa present. As a result, ranges of conodont species are known only in terms of trilobite zones, and the range of each conodont taxon within a specific trilobite zone is currently unknown unless it is present in the overlying and underlying zones. Müller and Hinz (Reference Müller and Hinz1991) established no conodont zones, but their study suggests that many taxa are long ranging, although some of their species have restricted stratigraphic ranges (Müller and Hinz, Reference Müller and Hinz1991, table 2). It seems likely that their classic monograph has contributed to the belief that many paraconodonts are stratigraphically long ranging and hence of limited biostratigraphic use. In addition, several other studies in Europe and elsewhere have been mainly general or taxonomic in nature, with relatively little attention paid to the potential use of especially paraconodonts as guide fossils (Müller, Reference Müller1959).
The fourth factor contributing to the lack of established Cambrian paraconodont biostratigraphy in most regions of the world is the lack of detailed research. Outside of China, there has been relatively little conodont research in strata older than the late late Cambrian, especially compared with the extensive work on euconodonts from younger rocks. For instance, many promising Cambrian sections in eastern and western North America remain unexplored for conodonts (Dong et al., Reference Dong, Bergström and Repetski2004a). For the past 25 years, the middle and lower upper Cambrian conodont biostratigraphic work in the world has largely been centered in China, and especially in South China.
The purpose of this paper is to describe conodonts of the upper Cambrian (Furongian Series) through lowermost Ordovician from Hunan, South China and to redescribe conodonts of the middle Cambrian, based on the material that has been accumulated for the past quarter century. The taxonomy of some conodont genera is revised. In light of histological investigation, some genera can now be assigned to euconodonts, paraconodonts, or protoconodonts.
Geological setting
In terms of the paleogeography of China, the Southeast Stratigraphic Region is characterized by flyschoid sediments. In the Yangtze Stratigraphic Region, the middle and upper Cambrian succession mainly is composed of dolostone. Only in the Langyashan Subregion does the middle and upper Cambrian consist mainly of dolomitic limestone. The upper part of the middle Cambrian through the lowermost Ordovician is made up chiefly of limestone in the Danzai-Baojing-Qinyang Subregion of the Jiangnan Stratigraphic Region. Within that area, mainly in western Hunan, are well-developed and continuously deposited stratigraphic successions. Accordingly, western Hunan is an ideal area for the study of Cambrian conodonts in South China.
Three sections in Hunan were selected as key sections. These are the Paibi section in Huayuan County, the Wangcun section in Yongshun County, and Wa’ergang section in Taoyuan County. Three other sections in Hunan were selected as auxiliary sections, including the Xiaoxi section in Jishou County, the Xiaoxiqiao section in Qianzhou, and the Yangjiazhai section in Jishou. Two additional sections in Hunan were selected as reference sections; these are the Tieqiao to Liangshuijing section in Fenghuang County and the Huaqiao section in Baojing County (Fig. 1). Some of the data gathered from these investigations have been published (Dong, Reference Dong1990a, Reference Dongb, Reference Dong1991, Reference Dong1992, Reference Dong1993, Reference Dong1997, Reference Dong1999a, Reference Dongb, Reference Dubinina2000, Reference Dong2001; Dong and Bergström, Reference Dong and Bergström2000a, Reference Dong and Bergströmb, Reference Dong and Bergström2001a, Reference Dong and Bergströmb; Dong et al., Reference Dong, Repetski and Bergström2001, Reference Dong, Repetski and Bergström2004a, Reference Dong, Bergström and Repetskic). The present study is the continuation of these previous studies and includes the first systematical description of the conodonts of upper Cambrian through lowermost Ordovician from Hunan, South China, taxonomic revision of some Cambrian conodonts, review of the problems in the zonation of paraconodonts recovered from western Hunan, South China, and assignment of some genera to euconodonts, paraconodonts, or protoconodonts in light of histological investigation.

Figure 1 Location of the studied sections: ☆ Location of the key sections, ▲ location of the auxiliary sections, △ location of the reference sections.
Conodont samples were collected from the three key sections on 15 occasions from 1986 to 2008 and from the three auxiliary sections from 1986 to 1995. The two reference sections were collected only sporadically, in 1986 and 1987. The average sampling interval is 2.5 m and the average sample mass is 4.6 kg. A total of 3,100 conodont samples have been collected, including re-sampled ones and 388 conodont-barren samples, with a total mass of more than 14,000 kg.
In the study area, there are mainly five types of lithology in the interval from the middle Cambrian Huaqiao Formation to the lower part of the lowermost Ordovician Panjiazui Formation.
-
1. Dark-gray, thin-bedded to medium- to thin-bedded, laminated, argillaceous, very fine to finely crystalline limy dolostone. Protoconodonts, but almost no other fossils, occur in this lithology.
-
2. Alternating layers of dark-gray, medium-bedded, laminated, argillaceous dolostone, and dark gray, medium-bedded micrite and biomicrite. The argillaceous dolostone does not yield fossils, whereas the limestones yield abundant conodonts, fossil embryos (Dong, Reference Dong2007a, Reference Dong2009a, Reference Dongb; Dong et al., Reference Dong, Donoghue, Cheng and Liu2004b, Reference Dong, Donoghue, Cunningham, Liu and Cheng2005a, Reference Dong, Bengtson, Gostling, Cunningham, Harvey, Kouchinsky, Val’kov, Repetski, Stampanoni, Marone and Donoghue2010; Peng and Dong, Reference Peng and Dong2008; Cheng et al., Reference Cheng, Peng, Duan and Dong2011), fossils of typical Orsten-type preservation (Dong et al., Reference Dong, Donoghue, Liu, Liu and Peng2005b; Liu and Dong, Reference Liu and Dong2007, Reference Liu and Dong2009, Reference Liu and Dong2010; Zhang and Dong, Reference Zhang and Dong2009; Zhang et al., Reference Zhang, Dong and Maas2011a, Reference Zhang, Dong and Xiaob, Reference Zhang, Dong and Xiao2012; Duan et al., Reference Duan, Dong and Donoghue2012), highly modified sponge spicules (Dong and Knoll, Reference Dong and Knoll1996; Chen and Dong, Reference Chen and Dong2008), radiolarians (Dong et al., Reference Dong, Knoll and Lipps1997), small shelly fossils (Zhu et al., Reference Zhu, Dong and Chen2003, Reference Zhu, Li and Dong2004; Zhu and Dong, Reference Zhu and Dong2004), as well as trilobites and brachiopods.
-
3. Dark-gray, thin- to medium-bedded biomicrosparite that yields abundant conodonts as well as some trilobites, brachiopods, and bradoriids.
-
4. Gray, thick-bedded calcirudite. Fragmentary trilobites have been found occasionally in this lithology. No conodont samples have been collected from this lithology.
-
5. Dark-gray, thin-bedded micrite intercalated with dark-gray, thin-bedded to laminated, dolomitic calcisiltite, containing organic matter and scattered pyrite. Burrows and bioturbation may occur. The micrite yields abundant conodonts, as well as trilobites, brachiopods, small shelly fossils, sponge spicules, and radiolarians.
Except for type 4, the calcirudites, the other four types of lithology were collected with specific sampling intervals from the key sections and auxiliary sections on the first sampling round. Mainly type 2, type 3, and type 5 were collected from the key sections during the second and third sampling rounds. Mainly type 3 and type 5 were collected during the fourth to the twelfth sampling rounds. The purpose of the fourth to the fifteenth sampling rounds was to recover, in addition to conodonts, sponge spicules, radiolarians, and soft-bodied fossils, such as embryos, and typical Orsten-type fossils, especially arthropods.
Materials and methods
The samples were processed by routine etching with ~10% technical acetic acid in plastic pails with a capacity of 10 L. The cycle of sieving and changing acid required 7–10 days. The samples (including duplicate samples) generally needed to be processed in three to four cycles for complete dissolution. During processing, the reaction time and the pH value of the solution were adjusted according to the lab temperature (~20°C in winter and up to 35°C in July and August). Some samples were processed in plastic pails with two-layered screens to recover better specimens for histologic studies as well as for recovery of phosphatized soft-bodied fossils. Results using the two-layered screens were very good.
For histological study, protoconodont-like elements were examined by double-polished thin sections and by etching both the thin sections and artificially fractured specimens with 0.5% orthophosphoric acid for 10 minutes, and then their images were taken by scanning electron microscope. Because of the small size and thin-walled nature of these paraconodonts and earliest euconodonts, the oil immersion technique is used for those elements (Dong et al., Reference Dong, Donoghue and Repetski2005c). Clove oil was used as the immersion medium, and specimens were examined using a Zeiss Axiophot or Olympus microscope fitted with differential interference contrast (Nomarski) optics. Photomicrographs were obtained using a Nikon Coolpix 990 or Olympus BH2 BHS UMA camera fitted via a c-mount and interchange lens to the microscope.
Repository and institutional abbreviation.—
All the figured specimens are deposited at the Geological Museum, Peking University (GMPKU).
Chronostratigraphy
The Ordovician is divided into Lower, Middle, and Upper Ordovician series. The current classification of Cambrian is divided into four series; Series 2 and Series 3 have not been formally named. The Terreneuvian and Series 2 are sometimes informally referred to as lower Cambrian, Series 3 is sometimes informally referred to as middle Cambrian, and the fourth series, the Furongian Series, is sometimes referred to as upper Cambrian. Therefore, we use the term Lower Ordovician formally, but also use the terms middle Cambrian and upper Cambrian informally.
Conodont biostratigraphy
Based on the stratigraphic ranges of conodont species, the middle Cambrian through lowermost Ordovician succession at Wangcun, Wa’ergang, and Paibi, South China, is subdivided into 13 conodont zones (Figs. 2–4; see also Dong and Bergström, Reference Dong and Bergström2001a).

Figure 2 Chart showing the stratigraphic ranges of the upper middle Cambrian through upper Cambrian (Furongian) conodonts and conodont zones in the Wangcun section in Hunan, China.

Figure 3 Chart showing the stratigraphic ranges of the upper middle Cambrian through lowermost Ordovician conodonts and conodont zones in the Wa’ergan section in Hunan, China.

Figure 4 Chart showing the stratigraphic ranges of the upper middle Cambrian conodonts and conodont zones in the Paibi section in Hunan, China (modified after Dong and Bergström, Reference Dong and Bergström2001a).
Gapparodus bisulcatus-Westergaardodina brevidens Zone.—
This zone is recognized in the lower part of the middle Cambrian Huaqiao Formation in the Paibi section, Huayuan and in the Wangcun section, Yongshun. Its lower limit is marked by the First Appearance Datum (FAD) of Gapparodus bisulcatus (Müller, Reference Müller1959), and its upper limit is defined by the FAD of Shandongodus priscus An, Reference An1982. The entire zone is characterized the dominance of protoconodonts and the first occurrence and high diversity of the genus Westergaardodina Müller, Reference Müller1959. The main taxa of the zone are Furnishina bigeminata Dong, Reference Dong1993, F. kleithria Müller and Hinz, Reference Müller and Hinz1991, G. bisulcatus, Gumella cuneata Müller and Hinz, Reference Müller and Hinz1991, Huayuanodontus tricornis (Dong, Reference Dong1993), Laiwugnathus hunanensis n. sp., Muellerodus? oelandicus (Müller, Reference Müller1959), Paibiconus proarcuatus Dong, Reference Dong1993, Phakelodus tenuis (Müller, Reference Müller1959), Westergaardodina brevidens Dong, Reference Dong1993, W. horizontalis Dong, Reference Dong1993, and Yongshunella polymorpha Dong and Bergström, 2001a.
Shandongodus priscus-Hunanognathus tricuspidatus Zone.—
This zone is recognized in the upper part of the middle Cambrian Huaqiao Formation in the Paibi and Wangcun sections. Its lower limit is defined by the FAD of Shandongodus priscus and its upper limit by the FAD of Westergaardodina quadrata (An, Reference An1982). It is characterized by the extreme diversity of paraconodonts and the maximum numbers of protoconodonts. Its main taxa are Coelocerodontus bicostatus van Wamel, Reference van Wamel1974, Furnishina cf. alata Szaniawski, Reference Szaniawski1971, F. cf. gladiata Müller and Hinz, Reference Müller and Hinz1991, F. cf. kranzae Müller and Hinz, Reference Müller and Hinz1991, F. longibasis Bednarczyk, Reference Bednarczyk1979, F. pernica An, Reference An1982, F. cf. quadrata Müller, Reference Müller1959, Hunanognathus tricuspidatus Dong, Reference Dong1993, Muellerodus? obliquus (An, Reference An1982), M. pomeranensis (Szaniawski, Reference Szaniawski1971), Nogamiconus sinensis (Nogami, Reference Nogami1966), S. priscus, Westergaardodina elegans Dong and Bergström, 2001a, W. tetragonia Dong, Reference Dong1993, and Westergaardodina sp. D. In addition, there are taxa ranging upward from the underlying zone, including F. bigeminata, Gapparodus bisulcatus, Gumella cuneata, Huayuanodontus tricornis, M.? oelandicus, Paibiconus proarcuatus, Phakelodus tenuis, W. brevidens, and Yongshunella polymorpha.
Westergaardodina quadrata Zone.—
This zone can be recognized in the lower part of the middle Cambrian Chefu Formation in the Paibi and Wangcun sections. Its lower limit is defined by the FAD of Westergaardodina quadrata and its upper limit by the FAD of W. matsushitai Nogami, Reference Nogami1966. It is characterized by the great abundance of W. quadrata. Its main taxa are Furnishina quadrata Müller, Reference Müller1959, Prosagittodontus compressus n. sp., W. cf. behrae Müller and Hinz, Reference Müller and Hinz1991, W. quadrata, W. cf. quadrata and W. sola n. sp. In addition, there are taxa ranging into this zone from the underlying zone, such as Furnishina cf. alata, F. bigeminata, F. kleithria, F. longibasis, F. pernica, F. cf. quadrata, Gapparodus bisulcatus, Gumella cuneata, Huayuanodontus tricornis, Hunanognathus tricuspidatus, Nogamiconus sinensis, Muellerodus? obliquus, M.? oelandicus, M. pomeranensis, Paibiconus proarcuatus, Phakelodus tenuis, W. elegans, W. tetragonia, and Yongshunella polymorpha.
Westergaardodina matsushitai-Westergaardodina grandidens Zone.—
This zone can be recognized in the upper part of the middle Cambrian Chefu Formation in the Paibi, Wangcun, and Wa’ergang sections. Its base is marked by the FAD of Westergaardodina matsushitai, and its top is defined by the FAD of Westergaardodina lui Dong, Repetski, and Bergström, 2004c. It is characterized by a notable reduction in the number of protoconodonts and by the occurrence of morphologically unusual species of Westergaardodina. Its main elements are Laiwugnathus transitans n. sp., L. cf. kouzhenensis An, Reference An1982, Prosagittodontus dahlmani (Müller, Reference Müller1959), Westergaardodina cf. horizontalis Dong, Reference Dong1993, W. gigantea n. sp., W. grandidens Dong, Reference Dong1993, and W. matsushitai. In addition, there are taxa ranging from the underlying zone, such as Coelocerodontus bicostatus, Furnishina cf. alata, F. bigeminata, F. cf. kranzae, F. longibasis, F. quadrata, F. cf. quadrata, Gapparodus bisulcatus, Gumella cuneata, Huayuanodontus tricornis, Muellerodus? obliquus, M.? oelandicus, M. pomeranensis, Nogamiconus sinensis, Paibiconus proarcuatus, Phakelodus tenuis, Prosagittodontus compressus n. sp., W. elegans, W. cf. quadrata, W. sola n. sp., W. quadrata and Yongshunella polymorpha.
Westergaardodina lui-Westergaardodina ani Zone.—
This zone occurs in the lower part of the upper Cambrian Bitiao Formation in the Wangcun and Wa’ergang sections. This zone supersedes the Westergaardodina proligula Zone as used in Dong and Bergström (Reference Dong and Bergström2001a). Its lower boundary is defined by the FAD of W. lui, and its top by the FAD of W. cf. calix Müller and Hinz, Reference Müller and Hinz1991. It is characterized by a notable reduction of the diversity of species of Westergaardodina compared with strata below. Its main taxa are Furnishina dayangchaensis Chen and Gong, Reference Chen and Gong1986, F. furnishi Müller, Reference Müller1959, F. cf. furnishi Müller, Reference Müller1959, F. gladiata Müller and Hinz, Reference Müller and Hinz1991, F. kranzae Müller and Hinz, Reference Müller and Hinz1991, F. primitiva Müller, Reference Müller1959, F. tortilis (Müller, Reference Müller1959), F. cf. ovata Müller and Hinz, Reference Müller and Hinz1991, Muellerodus cambricus (Müller, Reference Müller1959), M. guttulus Müller and Hinz, Reference Müller and Hinz1991, Proacodus obliquus Müller, Reference Müller1959, P. pulcherus (An, Reference An1982), Prooneotodus gallatini (Müller, Reference Müller1959), P. terashimai (Nogami, Reference Nogami1967), Prosagittodontus cf. eureka Müller, Reference Müller1959, Wangcunella conicus n. gen. n. sp., Wangcunognathus elegans n. gen. n. sp., Westergaardodina ahlbergi Müller and Hinz, Reference Müller and Hinz1991, W. ani Dong et al., Reference Dong, Donoghue, Cheng and Liu2004c, W. dimorpha n. sp., W. lui, W. microdentata Zhang in An et al., Reference An, Zhang, Xiang, Zhang, Xu, Zhang, Jiang, Yang, Lin, Cui and Yang1983, W. tricuspidata Müller, Reference Müller1959, and Westergaardodina sp. B. In addition, there are taxa that continue from the underlying zone, such as Coelocerodontus bicostatus, F. bigeminata, F. cf. kranzae, F. cf. gladiata, F. longibasis, F. quadrata, F. cf. quadrata, Gapparodus bisulcatus, Gumella cuneata, Huayuanodontus tricornis, Laiwugnathus transitans n. sp., M.? obliquus, M.? oelandicus, M. pomeranensis, Nogamiconus sinensis, Phakelodus tenuis, Prosagittodontus compressus n. sp., P. dahlmani, W. elegans, and Yongshunella polymorpha.
Westergaardodina cf. calix-Prooneotodus rotundatus Zone.—
This zone is recognized in the upper part of the upper Cambrian Bitiao Formation in the Wangcun and Wa’ergang sections. This zone supercedes the Westergaardodina cf. behrae-Prooneotodus rotundatus Zone as used in Dong and Bergström (Reference Dong and Bergström2001a). Its base is marked by the FAD of Westergaardodina cf. calix and its upper limit by the FAD of Proconodontus tenuiserratus Miller, Reference Miller1980. It is characterized by the maximum number of species of Prooneotodus observed in the various zones. Its main taxa are Furnishina wangcunensis n. sp., Granatodontus ani (Wang, Reference Wang1985), Hispidodontus resimus Nicoll and Shergold, Reference Nicoll and Shergold1991, Prooneotodus rotundatus (Druce and Jones, Reference Druce and Jones1971), Serratocambria minuta Müller and Hinz, Reference Müller and Hinz1991, Tujiagnathus gracilis n. gen. n. sp., W. cf. calix, W. wimani Szaniawski, Reference Szaniawski1971, Westergaardodina sp. A., and Westergaardodina sp. E. In addition, there are taxa ranging from the underlying zone, such as Coelocerodontus bicostatus, F. bigeminata, F. dayangchaensis, F. furnishi, F. cf. furnishi, F. kleithria, F. cf. kranzae, F. cf. gladiata, F. longibasis, F. cf. ovata, F. cf. quadrata, Huayuanodontus tricornis, Muellerodus cambricus, M.? oelandicus, M. pomeranensis, M.? obliquus, Phakelodus tenuis, Proacodus obliquus, P. pulcherus, Prooneotodus terashimai, Prosagittodontus dahlmani, Wangcunella conicus n. gen. n. sp., Westergaardodina ani, W. lui, and W. tricuspidata.
Proconodontus tenuiserratus Zone.—
This zone can be recognized in the lower part of the upper Cambrian Shengjiawan Formation in the Wa’ergang section. Its lower boundary is defined by the FAD of Proconodontus tenuiserratus and its upper boundary by the FAD of Proconodontus muelleri Miller, Reference Miller1969. Its main taxa are Coelocerodontus hunanensis n. sp., Millerodontus intermedius n. gen. n. sp., and P. tenuiserratus. In addition, there are taxa ranging from the underlying zone upward into this zone, such as Coelocerodontus bicostatus, Furnishina bigeminata, F. dayangchaensis, F. furnishi, Granatodontus ani, Huayuanodontus tricornis, Muellerodus cambricus, M.? oelandicus, Phakelodus tenuis, Proacodus obliquus, Prooneotodus gallatini, P. terashimai, P. rotundatus, Wangcunella conicus n. gen. n. sp., Westergaardodina ani, W. lui, and W. cf. calix.
Proconodontus Zone.—
This zone is recognized in the lower part of the upper Cambrian Shengjiawan Formation in the Wangcun and Wa’ergang sections. In addition, this zone can be identified in the upper Cambrian Cheshuitong Formation in Chuxian, Anhui (Dong, Reference Dong1987). Its lower and upper limits are defined by the FAD of Proconodontus muelleri and Eoconodontus notchpeakensis (Miller, Reference Miller1969), respectively. Its main taxa are Dasytodus transmutatus (Xu and Xiang in An et al., Reference An, Zhang, Xiang, Zhang, Xu, Zhang, Jiang, Yang, Lin, Cui and Yang1983), P. muelleri, P. posterocostatus Miller, Reference Miller1980, P. serratus Miller, Reference Miller1980, and Proacodus cf. pulcherus (An, Reference An1982). In addition, there are taxa that range from the underlying zone into this zone, such as Coelocerodontus bicostatus, C. hunanensis n. sp., Furnishina bigeminata, F. furnishi, F. cf. furnishi, F. dayangchaensis, F. cf. ovata, Granatodontus ani, Millerodontus intermedius n. gen. n. sp., Muellerodus cambricus, M.? obliquus, M.? oelandicus, Phakelodus tenuis, Proacodus obliquus, P. pulcherus, Proconodontus tenuiserratus, Prooneotodus gallatini, P. terashimai, P. rotundatus, Prosagittodontus dahlmani, Wangcunella conicus n. gen. n. sp., Westergaardodina ani, W. lui, and W. cf. calix.
Eoconodontus Zone.—
This zone can be recognized in the upper part of the upper Cambrian Shengjiawan Formation in the Wa’ergang section. Its lower and upper boundaries are defined by the FAD of Eoconodontus notchpeakensis and Cordylodus proavus, respectively. Its main taxa are E. notchpeakensis, Lugnathus hunanensis n. gen. n. sp., Hirsutodontus nodus (Zhang and Xiang, 1983), Mamillodus ruminates Dubinina, Reference Dubinina2000, Miaognathus multicostatus n. gen. n. sp., Prooneotodus? sp. A, Teridontus nakamurai (Nogami, Reference Nogami1967), Westergaardodina cf. nogamii Müller Hinz, Reference Müller and Hinz1991, and Westergaardodina sp. C. In addition, there are taxa ranging from the underlying zone into this zone, such as Coelocerodontus bicostatus, C. hunanensis n. sp., Furnishina bigeminata, F. dayangchaensis, F. furnishi, F. cf. furnishi, F. primitiva, F. cf. ovata, Dasytodus transmutatus, Granatodontus ani, Millerodontus intermedius n. gen. n. sp., Muellerodus cambricus, M.? oelandicus, Phakelodus tenuis, Proacodus obliquus, Prooneotodus gallatini, P. terashimai, P. rotundatus, Proconodontus muelleri, P. posterocostatus, P. serratus, Prosagittodontus dahlmani, Wangcunella conicus n. gen. n. sp., Westergaardodina cf. calix, W. ani, and W. lui.
Cordylodus proavus Zone.—
This zone is recognized in the upper part of the upper Cambrian Shengjiawan Formation in the Wa’ergang section. Moreover, this zone can be recognized in the upper Cambrian Cheshuitong Formation at Chuxian, Anhui (Dong, Reference Dong1987). Its lower and upper boundaries are defined by the FAD of Cordylodus proavus and C. intermedius Furnish, Reference Furnish1938, respectively. Its main taxa are C. proavus and Eodentatus bicuspatus Nicoll and Shergold, Reference Nicoll and Shergold1991. In addition, there are taxa ranging from the underlying zone, such as Coelocerodontus bicostatus, C. hunanensis n. sp., Dasytodus transmutatus, Eoconodontus notchpeakensis, Furnishina bigeminata, F. furnishi, F. dayangchaensis, F. primitiva, Lugnathus hunanensis n. gen. n. sp., Muellerodus? oelandicus, Phakelodus tenuis, Prooneotodus gallatini, P. rotundatus, P. terashimai, Prosagittodontus dahlmani, Teridontus nakamurai, Westergaardodina cf. calix, W. ani, and W. lui.
Cordylodus intermedius Zone.—
This zone is recognized in the lowermost part of the Panjiazui Formation in the Wa’ergang section. It can also be recognized in the lowermost part of the Cheshuitong Formation in Chuxian, Anhui (Dong, Reference Dong1987); that formation originally was assigned to the Lower Ordovician. Its lower boundary is marked by the FAD of Cordylodus intermedius, and its upper boundary by the FAD of C. lindstromi Druce and Jones, Reference Druce and Jones1971. Its main taxa are C. intermedius and C. prolindstromi Nicoll, Reference Nicoll1991. In addition, there are taxa ranging from the underlying zone, such as Coelocerodontus bicostatus, C. hunanensis n. sp., Cordylodus proavus, Eoconodontus notchpeakensis, Furnishina primitiva, Lugnathus hunanensis n. gen. n. sp., Muellerodus? oelandicus, Phakelodus tenuis, Prooneotodus gallatini, P. rotundatus, Teridontus nakamurai, Westergaardodina ani, W. cf. calix, and W. lui.
Cordylodus lindstromi Zone.—
This zone is recognized in the lower part of the Panjiazui Formation in the Wa’ergang section. Its lower and upper boundaries are defined by the FAD of Cordylodus lindstromi and C. angulatus, respectively. Its main taxa are C. caseyi Druce and Jones, Reference Druce and Jones1971, C. lindstromi, Iapetognathus fluctivagus Nicoll, Miller, Nowlan, Repetski, and Ethington, Reference Nicoll, Miller, Nowlan, Repetski and Ethington1999, I. jilinensis Nicoll, Miller, Nowlan, Repetski, and Ethington, Reference Nicoll, Miller, Nowlan, Repetski and Ethington1999, and Monocostodus sevierensis. Taxa that continue from the underlying zone include Coelocerodontus bicostatus, Cordylodus intermedius, C. proavus, C. prolindstromi, Eoconodontus notchpeakensis, Lugnathus hunanensis n. gen. n. sp., Phakelodus tenuis, Teridontus nakamurai, and Westergaardodina ani.
Cordylodus angulatus Zone (lower part).—
This zone is recognized in the lower part of the Panjiazui Formation in the Wa’ergang section. Its lower limit is defined by the FAD of Cordylodus angulatus, but its upper limit is not delimited herein because the upper portion of this zone is beyond the scope of the present study. Its main taxa are C. angulatus and Iapetognathus aengensis (Lindström, Reference Lindström1955). In addition, there are taxa that range from the underlying zone, such as C. intermedius, C. lindstromi, Monocostodus sevierensis, and Teridontus nakamurai.
Correlation of conodont zones
The formal conodont zonation older than the Fengshan Formation in North China and the Shengjiawan Formation in South China (Furongian, upper Cambrian) has been mainly established in China (Figs. 5, 6). Outside China, the middle Cambrian conodont zone Gapparodus bisulcatus-Westergaadodina mossebergensis is erected in the lower part of the Machari Formation, Yeongweol area, Gangweon Province, in South Korea (Lee et al., Reference Lee, Lee, Miller and Jeong2009).

Figure 5 Correlation of the upper middle Cambrian through lowermost Ordovician conodont zones in Hunan, South China, and those in North China, western USA, and Newfoundland, Canada.

Figure 6 Correlation between the conodont zones and trilobite zones of the upper middle Cambrian through lowermost Ordovician in Hunan, South China.
At present, the paraconodont zones below the Shengjiawan Formation in Hunan can be correlated only with those in North China (An, Reference An1982; Chen and Gong, Reference Chen and Gong1986). In contrast, eucondont zones in Hunan can be correlated widely with related zones in North China and elsewhere (Fig. 5).
Gapparodus bisulcatus-Westergaardodina brevidens Zone.—
In terms of species, this zone shares only a few elements with the Laiwugnathus laiwuensis Zone of North China (An, Reference An1982). The upper boundary of the Hunan zone and the L. laiwuensis Zone are both marked by the FAD of Shandongodus priscus, so the top of both zones may be correlated. In Hunan, the lowest occurrence of Gapparodus bisulcatus is in the lowermost part of the middle Cambrian Huaqiao Formation, and this level is equivalent to the lowest occurrence of the trilobite Ptychagnostus atavus (Tullberg, Reference Tullberg1880). The lowest occurrence of P. atavus in Hunan corresponds to that of Crepicephalina Resser and Endo in Kobayashi, 1935 in North China (Peng, Reference Peng2009a). Accordingly, it is inferred that the lower part of the G. bisulcatus-Westergaardodina brevidens Zone correlates with the lower part of the Zhangxia Formation in North China (Fig. 6).
Shandongodus priscus-Hunanognathus tricuspidatus Zone.—
This zone shares the zonal index species Shandongodus priscus with the S. priscus Zone in North China, so the two zones can be easily correlated with each other. The bases of the two zones are marked by the FAD of S. priscus, so the bases of these zones may be correlated. The top of the S. priscus-Hunanognathus tricuspidatus Zone in Hunan is marked by the FAD of Westergaardodina quadrata. W. quadrata and Westergaardodina orygma are associated in North China, and their FAD is at the same horizon. Accordingly, the top of the S. priscus-H. tricuspidatus Zone in Hunan appears to be identical to that of the S. priscus Zone in North China.
Westergaardodina quadrata Zone.—
Westergaardodina quadrata and W. orygma occur together and basically have the same range in the W. orygma Zone in North China. The base of both these zones precisely corresponds with each other. Because the tops of both zones are defined by the FAD of W. matsushitai, these zones represent the same stratigraphic interval, and the W. quadrata Zone in South China is equivalent to the W. orygma Zone in North China.
Westergaardodina matsushitai-Westergaardodina grandidens Zone.—
Westergaardodina matsushitai occurs in the W. matsushitai Zone in Hunan and in North China (An, Reference An1982). As mentioned above, the bases of both zones are defined in the same way and hence represent the same stratigraphic level. The top of the Hunan zone is defined by the FAD of W. lui, which is only a few meters higher than the last occurrence of W. grandidens and W. matsushitai, so it is nearly identical to the top of the latter zone. Therefore, the Hunan zone is correlated with the W. matsushitai Zone in North China.
Westergaardodina lui-Westergaardodina ani Zone.—
In Hunan, this zone shares only Proacodus pulcherus with the fauna of the Muellerodus? erectus Zone in North China (An, Reference An1982; Dong and Bergström, Reference Dong and Bergström2001a; Dong et al., 2004c). The base of the Hunan zone is defined by the FAD of Westergaardodina lui, and the base of the North China zone is defined by the last occurrence of W. matsushitai. In Hunan, the FAD of W. lui is only a few meters higher than the last occurrence of W. matsushitai and W. grandidens. Accordingly, the bases of both zones are roughly coeval. The top of the Hunan zone is marked by the FAD of W. cf. calix, whereas the top of the North China zone is defined as the level of the last appearance of M.? erectus. Precise correlation of the tops of these zones is difficult, and these zones are only approximately equivalent to each other.
Westergaardodina cf. calix-Prooneotodus rotundatus Zone.—
This zone shares Prooneotodus terashimai, and abundant P. gallatini with the Westergaardodina aff. fossa-P. rotundatus Zone in North China (An, Reference An1982; Dong and Bergström, Reference Dong and Bergström2001a; Dong et al., 2004c). The bases of both zones are broadly equivalent. The tops of both these zones are marked by the FAD of Proconodontus tenuiserratus, so these zones appear to represent the same stratigraphic interval.
Proconodontus tenuiserratus Zone.—
Proconodontus tenuiserratus occurs in the P. tenuiserratus Zone in Hunan, in the P. tenuiserratus Zone in North China (An, Reference An1982; Chen and Gong, Reference Chen and Gong1986), and in the western United States of America (Miller, Reference Miller1988; Miller et al., Reference Miller, Evens, Loch, Ethington, Stitt, Holmer and Popov2003). Because of the rather short stratigraphic range of this species, this zone in these three regions can be approximately correlated with one another.
Proconodontus Zone.—
Proconodontus muelleri and Teridontus nakamurai occur in the Proconodontus Zone in Hunan and in North China (An, Reference An1982; Chen and Gong, Reference Chen and Gong1986), and both taxa are also present in western USA (Miller, Reference Miller1988; Nicoll et al., Reference Nicoll, Miller, Nowlan, Repetski and Ethington1999; Miller et al., Reference Miller, Evens, Loch, Ethington, Stitt, Holmer and Popov2003). Because P. posterocostatus has been found only in two horizons in Hunan, the P. posterocostatus Subzone and the P. muelleri Subzone recognized in North China cannot be distinguished in Hunan. However, the Proconodontus Zone in Hunan roughly corresponds to the Proconodontus Zone in North China (An, Reference An1982; Chen and Gong, Reference Chen and Gong1986). The Hunan zone corresponds to the interval of both the P. posterocostatus Zone and the P. muelleri Zone recognized in the western USA (Miller, Reference Miller1988; Miller et al., Reference Miller, Evens, Loch, Ethington, Stitt, Holmer and Popov2003).
Eoconodontus Zone.—
Eoconodontus notchpeakensis occurs in the Eoconodontus Zone in Hunan and in the Cambrooistodus Zone of North China (Chen and Gong, Reference Chen and Gong1986) and in the Eoconodontus Zone in the western USA (Miller, Reference Miller1988; Miller et al., Reference Miller, Evens, Loch, Ethington, Stitt, Holmer and Popov2003, Reference Miller, Evens, Freeman, Ripperdan and Taylor2014a). In the western United States, the Eoconodontus Zone is subdivided into the E. notchpeakensis Subzone and the Cambrooistodus minutus Subzone. The correlation between these subzones and the Cambrooistodus Zone in North China (Chen and Gong, Reference Chen and Gong1986) is not clear because in North China the lowest part of the range of E. notchpeakensis is within the Proconodontus Zone (Chen and Gong, Reference Chen and Gong1986, text-fig. 26), and a separate Eoconodontus Zone is not recognized there. Cambrooistodus is not found in Hunan, and the Cambrooistodus Zone is only recognized at Dayangcha, North China in the warm (shallow) water realm (Miller, Reference Miller1988, fig. 3). The Eoconodontus Zone in Hunan is interpreted as broadly equivalent with the Eoconodontus Zone in the western United States (Miller, Reference Miller1988) and with the Cambrooistodus Zone of North China (Chen and Gong, Reference Chen and Gong1986). The Eoconodontus Zone in Hunan also roughly corresponds with the E. notchpeakensis Zone in western Newfoundland, Canada (Barnes, Reference Barnes1988), and with the Eoconodontus Zone in Black Mountain, western Queensland, Australia (Shergold et al., Reference Shergold, Nicoll, Laurie and Radke1991).
Cordylodus proavus Zone.—
Cordylodus proavus occurs in the C. proavus Zone in Hunan and in the C. proavus Zone in North China (An, Reference An1982; Chen and Gong, Reference Chen and Gong1986), and the two zones can be correlated with each other. The base of the zone is marked by the FAD of C. proavus and the top by the FAD of C. intermedius. The C. proavus Zone in Hunan is also correlated with the C. proavus Zone in the western USA (Miller, Reference Miller1980, Reference Miller1988; Nicoll et al., Reference Nicoll, Miller, Nowlan, Repetski and Ethington1999; Miller et al., Reference Miller, Evens, Loch, Ethington, Stitt, Holmer and Popov2003), and with the interval containing the proavus Fauna in western Newfoundland (Barnes, Reference Barnes1988; Cooper et al., Reference Cooper, Nowlan and Williams2001).
Cordylodus intermedius Zone.—
This zone is recognized worldwide. In the western United States, according to Miller (Reference Miller1988), “the base of the Cordylodus intermedius Zone is at the lowest occurrence of several euconodonts (Hirsutodontus simplex, Monocostodus sevierensis, Semiacontiodus lavadamensis, Utahconus utahensis, and C. intermedius), and the paraconodont Albiconus postcostatus Miller, Reference Miller1980. In some sections, all of these species begin at the same level or if one occurs below the others, it is often H. simplex.” The C. intermedius Zone is divided into the H. simplex and Clavohamulus hintzei Subzones in the western United States. In the Dayangcha sections in northeastern China, the first appearance of C. intermedius, H. simplex, U. utahensis, S. lavadamensis, M. sevierensis, and A. postcostatus is at the same, or nearly the same, horizon, so the base of the C. intermedius Zone there correlates with the base of the H. simplex Subzone of the North American zonal scheme (Chen and Gong, Reference Chen and Gong1986). However, H. simplex and C. hintzei have not yet been found in Hunan, probably because these two species were shallow water taxa. Rare specimens of these species have been found at the Green Point section, Newfoundland, but these specimens may be reworked from shallow water platform strata (Cooper et al., Reference Cooper, Nowlan and Williams2001). The precise correlation of the C. intermedius Zone in Hunan with the C. intermedius Zone in other regions is somewhat unclear. This is particularly the case with correlation of the top of the C. intermedius Zone in Hunan with that of the C. intermedius zones in other regions. Therefore, we interpret the C. intermedius Zone in Hunan as only roughly equivalent to the C. intermedius Zone in North China (Chen and Gong, Reference Chen and Gong1986), the C. intermedius Zone in the western United States (Miller, Reference Miller1988; Nicoll et al., Reference Nicoll, Miller, Nowlan, Repetski and Ethington1999; Miller et al., Reference Miller, Evens, Loch, Ethington, Stitt, Holmer and Popov2003), and the interval of the intermedius Fauna in western Newfoundland (Cooper et al., Reference Cooper, Nowlan and Williams2001).
Cordylodus lindstromi Zone.—
This zone is recognized widely in the world. We tentatively consider the Cordylodus lindstromi Zone in Hunan as broadly corresponding to the C. lindstromi Zone in North China (Chen and Gong, Reference Chen and Gong1986). The Hunan zone corresponds to the combined Iapetognathus Zone and the Cordylodus lindstromi Zone in the western USA (Ross et al., Reference Ross, Hintze, Ethington, Miller, Taylor and Repetski1997; Nicoll et al., Reference Nicoll, Miller, Nowlan, Repetski and Ethington1999; Miller et al., Reference Miller, Evens, Loch, Ethington, Stitt, Holmer and Popov2003), and roughly to the Iapetognathus fluctivagus Zone (lindstromi-prion-Iapetognathus Fauna) in western Newfoundland (Cooper et al., Reference Cooper, Nowlan and Williams2001). Terfelt et al. (Reference Terfelt, Bagnoli and Stouge2012) restudied the Global boundary Stratotype Section and Point (GSSP) for the base of the Ordovician System in the Green Point section, Newfoundland, Canada. They found I. fluctivagus is not present at the boundary interval. As a result, the identification of the Cambrian–Ordovician boundary level outside the GSSP section has proven to be problematic. Miller et al. (Reference Miller, Evens, Loch, Ethington, Stitt, Holmer and Popov2003, Reference Miller, Repetski, Nicoll, Nowlan and Ethington2014b) suggested the base of Iapetognathus Zone should be the Cambrian–Ordovician boundary level, i.e., the FAD of lowest species of Iapetognathus. In Hunan, I. jilinensis Nicoll et al., Reference Nicoll, Miller, Nowlan, Repetski and Ethington1999 is the lowest species of Iapetognathus. Consequently, the FAD of I. jilinensis is the base of the upper part of the C. lindstromi Zone and the Cambrian–Ordovician boundary level. Unfortunately, I. jilinensis has been found at only one level in Hunan, so this correlation is only approximate.
Cordylodus angulatus Zone (lower part).—
Cordylodus angulatus occurs in the C. angulatus Zone in Hunan and in the C. angulatus-Chosonodina herfurthi Zone in North China (Chen and Gong, Reference Chen and Gong1986), in the C. angulatus Zone in the western United States (Miller, Reference Miller1980, Reference Miller1988; Ross et al., Reference Ross, Hintze, Ethington, Miller, Taylor and Repetski1997; Nicoll et al., Reference Nicoll, Miller, Nowlan, Repetski and Ethington1999; Miller et al., Reference Miller, Evens, Loch, Ethington, Stitt, Holmer and Popov2003), and in the interval of the angulatus Fauna in western Newfoundland (Barnes, Reference Barnes1988; Cooper et al., Reference Cooper, Nowlan and Williams2001). It appears that the C. angulatus Zone in Hunan corresponds to the lower part of the zone in the regions mentioned, and the top of this zone is above the stratigraphic interval studied herein.
Correlation of conodont zones with trilobite zones
Many workers have investigated the trilobite biostratigraphy of Cambrian sections in Hunan, e.g., Lu (Reference Lu1956), Jegorova et al. (Reference Jegorova, Xiang, Lee, Nan and Kuo1963), Lin et al. (Reference Lin, Wang and Liu1966), Yang (Reference Yang1978, Reference Yang1981, Reference Yang1984), Lin (Reference Lin1991), Dong (Reference Dong1990a, Reference Dongb, 1991), Peng (Reference Peng1987, Reference Peng1992), Peng and Robison (Reference Peng and Robison2000), Peng et al. (Reference Peng, Babcock, Robison, Lin, Rees and Saltzman2004), Peng (Reference Peng2009a, Reference Pengb) and Peng et al. (Reference Peng, Babcock and Cooper2012). Peng (Reference Peng2009a, Reference Pengb) and Peng et al. (Reference Peng, Babcock and Cooper2012) presented a comprehensive summary with much new information that allows detailed correlation with the Cambrian conodont zones in Hunan (Fig. 6).
Conodont histology
Although histological study on conodonts can be traced back to the 1930s (Branson and Mehl, Reference Branson and Mehl1933), systematic histological investigations on conodonts began in the 1970s (Müller and Nogami, Reference Müller and Nogami1971) and since then has been carried out by many authors (Bengtson, Reference Bengtson1976, Reference Bengtson1983; Szaniawski, Reference Szaniawski1982, 1983, 1987; Andres, Reference Andres1988; Szaniawski and Bengtson, Reference Szaniawski and Bengtson1993, Reference Szaniawski and Bengtson1998; Müller and Hinz, Reference Müller and Hinz1998, etc.). The discovery of the entire conodont animal (Briggs et al., Reference Briggs, Clarkson and Aldridge1983; Aldridge, Reference Aldridge1987; Aldridge et al., Reference Aldridge, Briggs, Smith, Clarkson and Clark1993; Gabbott et al., Reference Gabbott, Aldridge and Theron1995) greatly stimulated the comparative histological study on euconodonts so as to verify their biological affinity, and many papers were involved in the “conodont controversies” (Purnell et al., Reference Purnell, Aldridge, Donoghue and Gabbot1995; Aldridge and Purnell, Reference Aldridge and Purnell1996; Donoghue et al., Reference Donoghue, Forey and Aldridge2000). In contrast to the debate, Szaniawski did refined work on the affinity of protoconodonts based on their hard tissues and the fossils of chaetognaths recognized in the Burgess Shale (Szaniawski, Reference Szaniawski2002, Reference Szaniawski2005). Philip C. J. Donoghue and Xi-ping Dong investigated well-preserved elements of protoconodonts, paraconodonts, and the earliest euconodonts from the middle and late Cambrian in the Paibi, Wangcun, and Wa’ergang sections in werstern Hunan. They found that conodont elements recovered from western Hunan are of great significance in terms of histology. Our comparative histological study on the earliest euconodonts has been published (Dong et al., Reference Dong, Donoghue and Repetski2005c; Guo et al., Reference Guo, Dong, Zeng and Zhao2005a, Reference Guo, Dong, Zeng and Zhaob), and the study on paraconodonts will be published in a separate paper.
Assignment of taxa to either protoconodonts, paraconodonts, or euconodonts is vitally important in the study of Cambrian conodonts. The present paper is only involved in the histological study as it relates to assigning taxa to one of those three groups. Bengtson (Reference Bengtson1976) proposed a model showing an evolutionary sequence from protoconodonts through paraconodonts to euconodonts. A direct evolutionary relationship between protoconodonts and paraconodonts remains unproved and is much more hypothetical than that between paraconodonts and euconodonts (Szaniawski and Bengtson, Reference Szaniawski and Bengtson1993; Donoghue et al., Reference Donoghue, Forey and Aldridge2000; Murdock et al., Reference Murdock, Dong, Repetski, Marone, Stampanon and Donoghue2013). At present, protoconodonts are assigned to phylum Chaetognatha Leuckart, Reference Leuckart1854, whereas paraconodonts and euconodonts are assigned to different classes of phylum Chordata Bateson, Reference Bateson1886 (Lee et al., Reference Lee, Lee, Miller and Jeong2009).
Histological investigations by Philip C. J. Donoghue and Xi-ping Dong (unpublished data) indicate that it is not always possible to distinguish reliably among the elements of these three taxonomic groups based only on study of their external morphology, even by experienced observers, and histological study is often required to determine the correct taxonomy. Paibiconus Dong, Reference Dong1993 and Huayuanodontus Dong and Bergström, 2001a were both considered originally to be protoconodonts (An and Mei, Reference An and Mei1994; Dong and Bergström, 2001a). Histological investigation by Philip C. J. Donoghue and Xi-ping Dong verified that Paibiconus is a protoconodont (Dong, Reference Dong2004). However, longitudinal cross sectioning of elements of Huayuanodontus showed a two-layered structure that is different from that of protoconodonts, paraconodonts, or euconodonts. Therefore, further histological work is needed to determine the taxonomic assignment of Huayuanodontus (cf. Dong, Reference Dong2007b). Coelocerodontus Ethington, Reference Ethington1959 has thin-walled coniform elements with large basal cavities. The cross section is extremely narrow. Clusters are very common. It has been referred to as a protoconodont, paraconodont, or euconodont by different authors (Müller and Hinz, Reference Müller and Hinz1991, p. 53). Müller and Hinz (Reference Müller and Hinz1991) considered it to be a euconodont, although they did not elucidate their histological evidence in 1991 or later (Müller and Hinz, Reference Müller and Hinz1998). Philip C. J. Donoghue and Xi-ping Dong tested many specimens of this genus recovered from Hunan by means of immersion in clove oil (unpublished data). Unequivocal crown material was found in all the tested specimens, so this genus should be considered as a euconodont.
Prooneotodus rotundatus is a cosmopolitan paraconodont species. Miller (Reference Miller1980) recognized a new genus and species that resembles P. rotundatus. He stated that the new genus and species differs from P. rotundatus in possessing a thin layer of apatite (crown material) that covers a massive white basal cone (Miller, Reference Miller1980, p. 32). Unfortunately, he has not performed any histological investigations that he mentioned would be needed to confirm his interpretation (personal communication, J.F. Miller, 2007). We investigated well-preserved elements that appear to be P. rotundatus from upper Cambrian strata in Hunan, South China by means of the oil immersion technique (Dong et al., Reference Dong, Donoghue and Repetski2005c). Some of the investigated elements show typical paraconodont strucure. However, some investigated elements have crown tissues, in which we found crystals of apatite perpendicular to the conodont grown axis, and regeneration could be observed at the apical portion of the elements. Both features are typical of euconodonts, and these elements are considered euconodonts rather than paraconodonts. Herein, they are assigned to Millerodontus intermedius n. gen. n. sp., and the species is interpreted as a transition between the paraconodont Prooneotodus and the primitive euconodont genus Proconodontus. Regenerated tips of elements are found also in Proconodontus, which occurs with Millerodontus. This is the first report to establish a new conodont genus in the light of histological study.
Post-Tremadocian conodonts are all euconodonts, but protoconodonts, paraconodonts, and euconodonts co-occur in Cambrian through Tremadocian strata. Therefore, histological investigation of Cambrian conodonts is of great importance not only in the differentiation of protoconodonts, paraconodonts, and euconodonts, but this information is the foundation on which to explore the taxonomy and the early evolution of conodonts.
Systematic paleontology
Recently, Lee et al. (Reference Lee, Lee, Miller and Jeong2009) separated the Cambrian and Tremadocian conodnts into three groups: paraconodonts, euconodonts, and protoconodonts in their Systematic Paleontology and proposed an updated classification of conodont faunas in terms of phylum, class, and order. We follow their taxonomic treatment herein.
Euconodonts
Phylum Chordata Bateson, Reference Bateson1886
Superclass Conodonta Pander, Reference Pander1856
Class Conodonti Branson, Reference Branson1938
Genus Coelocerodontus Ethington, Reference Ethington1959
Type species.—
Coelocerodontus trigonius Ethington, Reference Ethington1959.
Remarks.—
This genus consists of thin-walled coniform elements with large basal cavities. The cross section is extremely narrow. Clusters are very common. As mentioned above, histological investigation indicates that it is a euconodont. It is probably the oldest known euconodont.
Coelocerodontus bicostatus van Wamel, Reference van Wamel1974
Figures 7.1–7.12, 7.16–7.18, 8.1–8.4

Figure 7 (1–12, 16–18) Coelocerodontus bicostatus van Wamel, Reference van Wamel1974; (1) Shenjiawan Formation, Wangcun section, GMPKU2399, lateral view; (2) Bitiao Formation, Wangcun section, GMPKU2400, lateral view; (3) Bitiao Formation, Wangcun section, GMPKU2401, lateral view; (4) Bitiao Formation, Wangcun section, GMPKU2402, lateral view; (5) Bitiao Formation, Wangcun section, GMPKU2403, lateral view; (6) Shenjiawan Formation, Wa’ergang section, GMPKU2404, lateral view; (7) Bitiao Formation, Wa’ergang section, GMPKU2405, lateral view; (8) Huaqiao Formation, Wangcun section, GMPKU2413, (9) Bitiao Formation, Wa’ergang section, GMPKU2406, lateral view; (10) Shenjiawan Formation, Wa’ergang section, GMPKU2407, lateral view; (11) Panjiazui Formation, Wa’ergang section, GMPKU2408, lateral view; (12) Bitiao Formation, Wa’ergang section, GMPKU2409, lateral view; (16) Panjiazui Formation, Wa’ergang section, GMPKU2410, lateral view; (17) Panjiazui Formation, Wa’ergang section, GMPKU2411, lateral view; (18) Panjiazui Formation, Wa’ergang section, GMPKU2412, lateral view; (13–15, 19–25) Coelocerodontus hunanensis n. sp.; (13) Shenjiawan Formation, Wangcun section, GMPKU2414, lateral view; (14) Shenjiawan Formation, Wangcun section, GMPKU2415, lateral view; (15) Shenjiawan Formation, Wa’ergang section, GMPKU2416, cluster; (19) Shenjiawan Formation, Wa’ergang section, GMPKU2417, lateral view; (20) Shenjiawan Formation, Wa’ergang section, GMPKU2418, lateral view; (21) Shenjiawan Formation, Wa’ergang section, GMPKU2419, lateral view; (22) Shenjiawan Formation, Wa’ergang section, GMPKU2420, lateral view; (23) Shenjiawan Formation, Wa’ergang section, GMPKU2421, lateral view; (24) Shenjiawan Formation, Wa’ergang section, GMPKU2422, lateral view; (25) Shenjiawan Formation, Wa’ergang section, GMPKU2423, lateral view. Relative scale bar represents 147 μm (1–11, 14, 16–25), 135 μm (12, 13, 15).

Figure 8 Conodont images using oil immersion techniques (Dong et al., 2005c) with Differential Interference Contrast (Nomarski) illumination. (1–4) Coelocerodontus bicostatus van Wamel, Reference van Wamel1974; (1) overview, GMPKU2254, showing the crown structure of euconodont; (2) close-up of (1); (3) overview, GMPKU2255, showing the crown structure of euconodont; (4) close-up of (3). Relative scale bar represents 109 μm (1, 3), 38 μm (2, 4).
1974Coelocerodontus bicostatus Reference van Wamelvan Wamel, p. 55, pl. 3, fig. 2.
1974Coelocerodontus latus Reference van Wamelvan Wamel, p. 56, pl. 1, fig. 2.
1983“Coelocerodontus” bicostatus van Wamel; Reference LandingLanding, p. 1172, fig. 10A, B.
1986Stenodontus compressus Reference Chen and GongChen and Gong, p. 186, pl. 24, figs. 11, 16, pl. 25, figs. 2–5, 7–13, 16, text-fig. 76.
1986Stenodontus jilinensis Reference Chen and GongChen and Gong, p. 187, pl. 18, figs. 2, 4–7, 9, 17, 18, pl. 19, figs. 3, 7, pl. 24, figs. 1, 9, 18, pl. 34, figs. 9, 14, 15, 19, text-fig. 77.
1987Coelocerodontus bicostatus van Wamel; Reference AnAn, p. 104, pl. 1, fig. 11.
1987Diaphanodus latus (van Wamel); Bagnoli et al., p. 155, pl. 2, figs. 11, 12.
1988Coelocerodontus cambricus (Nogami); Reference Heredia and BordonaroHeredia and Bordonaro, p. 190, pl. 2, figs. 2, 3.
1988Rotundoconus mendozanus Reference Heredia and BordonaroHeredia and Bordonaro, p. 194, pl. 3, fig. 5, pl. 4, fig. 3.
1988Coelocerodontus cambricus (Nogami); Reference Lee and LeeLee and Lee, pl. 1, fig. 27.
1988Coelocerodontus apparatus, Andres, pl. 9, figs. 3–8, text-fig. 19.
1991Coelocerodontus bicostatus van Wamel; Reference Müller and HinzMüller and Hinz, p. 53, pl. 41, figs. 1–21, text-figs. 20A–D.
2000“Proacontiodus” latus entis Reference Pyle and BarnesDubinina, p. 175, pl. 3, figs. 15, 16, 20, 21 (not figs. 1, 1a, 7, 8).
2000“Proacontiodus”latus Reference Pyle and BarnesDubinina, p. 174, pl. 3, figs. 30, 31.
2001Coelocerodontus cambricus (Nogami); Reference LeeLee, p. 449, figs. 6, 11.
2002aCoelocerodontus cambricus (Nogami); Reference LeeLee, p. 165, pl. 2, fig. 16.
2002bCoelocerodontus cambricus (Nogami); Reference LeeLee, p. 25, pl. 1, figs. 16, 17.
? 2006Coelocerodontus bicostatus van Wamel; Reference Qi, Bagnoli and WangQi et al., p. 187, pl. 3, fig. 17.
2007Coelocerodontus bicostatus van Wamel; Reference Landing, Westrop and KeppieLanding et al., p. 922, fig. 9a, b.
2009Coelocerodontus bicostatus van Wamel; Reference Lee, Lee, Miller and JeongLee et al., p. 423, figs. 7, 18.
2011Coelocerodontus bicostatus van Wamel; Reference Bagnoli and QiBagnoli and Qi, p. 12 (G, H, I).
2014Stenodontus compressus Chen and Gong; Reference Bagnoli and StougeBagnoli and Stouge, p. 20, fig. 11, N–R.
Description.—
Bimembrate apparatus of asymmetrical and subsymmetrical elements. The asymmetrical element is thin-walled and coniform. It is laterally compressed, with keeled anterior and posterior edges. Its cross section is extremely narrow. Both lateral sides have a costa that extends up to the apex. The pointed apex extends far beyond the posterior basal margin, leading to a recurved triangular outline in lateral view. The basal cavity is very deep. The subsymmetrical element differs from the asymmetrical element in the hook-like apical portion and the inconspicuous carinae instead of costae on both lateral sides.
Materials.—
469 specimens.
Occurrence.—
Known from the Wangcun and Wa’ergang sections, where it ranges from the Shandongodus priscus-Hunanognathus tricuspidatus Zone through the Cordylodus intermedius Zone.
Remarks.—
Müller and Hinz (Reference Müller and Hinz1991) described Morphotype alpha and Morphotype beta of this species. Of the material at hand, the asymmetrical element and subsymmetrical element correspond, respectively, with Morphotype alpha and Morphotype beta in terms of overall morphology.
Coelocerodontus hunanensis new species
Figures 7.13–7.15, 7.19–7.25, 9.1–9.14, 9.18, 9.19

Figure 9 (1–14, 18, 19) Coelocerodontus hunanensis n. sp.; (1) Shenjiawan Formation, Wa’ergang section, GMPKU2424, lateral view; (2) Shenjiawan Formation, Wa’ergang section, GMPKU2425, lateral view; (3) Shenjiawan Formation, Wa’ergang section, GMPKU2426, cluster, lateral view; (4) Shenjiawan Formation, Wa’ergang section, GMPKU2427, lateral view; (5) Shenjiawan Formation, Wa’ergang section, GMPKU2428, lateral view; (6) Shenjiawan Formation, Wa’ergang section, GMPKU2429, cluster, lateral view; (7) Panjiazui Formation, Wa’ergang section, GMPKU2430, cluster, lateral view; (8) Panjiazui Formation, Wa’ergang section, GMPKU2431, lateral view; (9) Panjiazui Formation, Wa’ergang section, holotype, GMPKU2432, lateral view; (10) Panjiazui Formation, Wa’ergang section, GMPKU2433, lateral view; (11) Panjiazui Formation, Wa’ergang section, GMPKU2434, cluster, lateral view; (12) Panjiazui Formation, Wa’ergang section, GMPKU2435, lateral view; (13) Panjiazui Formation, Wa’ergang section, GMPKU2436, cluster, lateral view; (14) Panjiazui Formation, Wa’ergang section, GMPKU2437, cluster, lateral view; (18) Panjiazui Formation, Wa’ergang section, GMPKU2438, lateral view; (19) Panjiazui Formation, Wa’ergang section, GMPKU2439, lateral view; (15, 17, 20, 22–24) Cordylodus angulatus Pander, Reference Pander1856; (15) Panjiazui Formation, Wa’ergang section, GMPKU2440, lateral view; (17) Panjiazui Formation, Wa’ergang section, GMPKU2441, lateral view; (20) Panjiazui Formation, Wa’ergang section, GMPKU2442, lateral view; (22) Panjiazui Formation, Wa’ergang section, GMPKU2169, lateral view; (23) Panjiazui Formation, Wa’ergang section, GMPKU2170, lateral view; (24) Panjiazui Formation, Wa’ergang section, GMPKU2171, lateral view; (16) Cordylodus proavus Müller, Reference Müller1959, Panjiazui Formation, Wa’ergang section, GMPKU2443, lateral view; (21, 25, 26) Cordylodus caseyi Druce and Jones, Reference Druce and Jones1971; (21) Panjiazui Formation, Wa’ergang section, GMPKU2444, lateral view; (25) Panjiazui Formation, Wa’ergang section, GMPKU2446, lateral view; (26) Panjiazui Formation, Wa’ergang section, GMPKU2447, posterior view. Relative scale bar represents 158 μm (1, 2, 4–19, 22), 150 μm (3, 20, 21, 23–26).
Diagnosis.—
Elongate proclined, thin-walled coniform elements with very deep basal cavities, laterally compressed, with keeled anterior and posterior edges. Both lateral sides are characterized by costae or only one lateral side is characterized by a costa. The outline is extremely slender, with a high length/maximum width ratio.
Description.—
Elongate, thin-walled coniform elements, proclined, asymmetrical, laterally compressed, with keeled anterior and posterior edges. The cross section is extremely narrow. Both lateral sides are characterized by costae or only one lateral side is characterized by a costa, each of which extends up to the apex; position of these costae is highly variable. The outline of the element is extremely slender. The length/maximum width ratio is more than 3.5. The tip of the cusp is usually not preserved. The basal cavity is very deep.
Etymology.—
Named for its provenance in Hunan Province.
Types.—
Holotype: GMPKU2432, from the Cordylodus intermedius Zone, Furongian (upper Cambrian), Panjiazui Formation, Wa’ergang section, Wa’ergang village, Taoyuan County, Hunan Province.
Materials.—
368 specimens.
Occurrence.—
Known from Wangcun and Wa’ergang sections, where it ranges from the Proconodontus tenuiserratus Zone through the Cordylodus intermedius Zone.
Remarks.—
It differs from Coelocerodontus bicostatus in its much more slender outline and much higher length/maximum width ratio.
Genus Cordylodus Pander, Reference Pander1856
Type species.—
Cordylodus angulatus Pander, Reference Pander1856.
Remarks.—
Most species of this genus are cosmopolitan in distribution and are of great significance in biostratigraphy and the evolution of early euconodonts. There has never been a consensus on their apparatus reconstruction. Miller (Reference Miller1980) established a two-element apparatus of this genus, and he identified them as rounded and compressed elements based on the cross section of the cusp. This interpretation was accepted by some authors based on their own collections, e.g., An et al. (Reference An, Zhang, Xiang, Zhang, Xu, Zhang, Jiang, Yang, Lin, Cui and Yang1983), Chen and Gong (Reference Chen and Gong1986), and Dong (Reference Dong1987) (only Cordylodus proavus). Viira et al. (Reference Viira, Sergeeva and Popovu1987) recognized three element types of C. proavus, whereas Barnes (Reference Barnes1988) suggested four element types. Nicoll (Reference Nicoll1990) reinterpreted the apparatus reconstruction of this genus as a septimembrate apparatus containing M, S, and P elements. We still use the apparatus reconstruction by Miller (Reference Miller1980). Although this two-element model does not seem to be refined and updated, it is more practical than more complex models. Miller’s rounded element includes Nicoll’s S elements and probably Nicoll’s P elements, and Miller’s compressed elements correspond to Nicoll’s M elements.
Cordylodus angulatus Pander, Reference Pander1856
Figures 9.15, 9.17, 9.20, 9.22–9.24, 10.1, 10.2

Figure 10 Conodont images using oil immersion techniques (Dong et al., 2005c) with Differential Interference Contrast (Nomarski) illumination, showing the shapes of basal cavities. (1, 2) Cordylodus angulatus Pander, Reference Pander1856; (1) the same specimen as Figure 9.17; (2) the same specimen as Figure 9.20; (3–6) Cordylodus intermedius Furnish, Reference Furnish1938; (3) the same specimen as Figure 12.1; (4) the same specimen as Figure 12.2; (5) the same specimen as Figure 12.3; (6) the same specimen as Figure 12.7. Relative scale bar represents 167 μm (1), 109 μm (2, 5), 103 μm (3, 4, 6).
1856Cordylodus angulatus Reference PanderPander, p. 33, pl. 2, figs. 28–31, pl. 3, fig. 10.
1856Cordylodus rotundatus Reference PanderPander, p. 33, pl. 2, figs. 32, 33.
1938Cordylodus subangulatus Reference FurnishFurnish, p. 338, text-fig. 2D, pl. 42, fig. 3.
1955Cordylodus angulatus Pander; Reference LindströmLindström, p. 551, pl. 5, fig. 9, text-fig. 3 (G).
1955Cordylodus rotundatus Pander; Reference LindströmLindström, p. 553, pl. 5, figs. 17–20, text-fig. 3 (F).
1971Cordylodus angulatus Pander; Reference Druce and JonesDruce and Jones, p. 66, pl. 3, figs. 4–6, text-figs. 23a, b.
1971Cordylodus rotundatus Pander; Reference Druce and JonesDruce and Jones, p. 71, text-figs. 23t, pl. 3, figs. 8–10.
1971Cordylodus sp. A Reference Druce and JonesDruce and Jones, p. 72, text-fig. 23u, pl. 8, fig. 10a, b.
1971Cordylodus rotundatus Pander; Reference Ethington and ClarkEthington and Clark, pl. 1, fig. 17.
1971Cordylodus angulatus Pander; Reference Ethington and ClarkEthington and Clark, pl. 1, 15, 16, 20.
1973Cordylodus angulatus Pander; Reference MüllerMüller, p. 27, pl. 11, figs. 1–7, text-fig. 2G.
1973Cordylodus rotundatus Pander; Reference MüllerMüller, p. 36, text-figs. 2H, 10a, b, pl. 11, figs. 8–10.
1974Cordylodus rotundatus Pander; Reference van Wamelvan Wamel, pl. 1, fig. 14.
1980Cordylodus angulatus Pander; Reference MillerMiller, p. 13, pl. 1, figs. 22, text-fig. 4Q.
1980Cordylodus rotundatus Pander; Reference MillerMiller, p. 20–21, pl. 1, fig. 24, text-fig. 4p.
1981Cordylodus angulatus Pander; Reference AnAn, pl. 2, fig. 17.
1981Cordylodus rotundatus Pander; Reference AnAn, pl. 1, figs. 18–19.
1982Cordylodus angulatus Pander; Reference RepetskiRepetski, p. 16, pl. 4, fig. 9, text-fig. 4 (L).
1982Cordylodus rotundatus Pander; Reference RepetskiRepetski, p. 18, pl. 5, fig. 3, text-fig. 4 (N).
1983Cordylodus angulatus Pander; Reference An, Zhang, Xiang, Zhang, Xu, Zhang, Jiang, Yang, Lin, Cui and YangAn et al., p. 84, pl. 8, figs. 1, 2.
1983Cordylodus rotundatus Pander; Reference An, Zhang, Xiang, Zhang, Xu, Zhang, Jiang, Yang, Lin, Cui and YangAn et al., p. 88–89, pl. 8, figs. 3–7.
1985Cordylodus rotundatus Pander; Reference NowlanNowlan, p. 111, text-fig. 4 (3).
1985Cordylodus angulatus Pander; Reference WangWang, p. 215, pl. 1, figs. 1–3, pl. 7, figs. 11–14, pl. 10, figs. 17 (not 15), pl. 11, figs. 17, 18 (?), pl. 12, figs. 18 (?), 19, 20 (?), pl. 14, fig. 7 (not 8) (part).
1985Cordylodus rotundatus Pander; Reference WangWang, p. 219, pl. 1, fig. 5, pl. 7, figs. 25, 26; pl. 11, fig. 22.
1985Cordylodus angulatus Pander; Reference DongDong, p. 394, pl. 1, fig. 6, pl. 3, fig. 5, text-fig. 1 (J).
1985Cordylodus rotundatus Pander; Reference DongDong, p. 396, pl. 2, fig. 8, pl. 3, fig. 9, text-fig. 1L.
1986Cordylodus angulatus Pander; Reference Chen and GongChen and Gong, p. 125, pl. 34, figs. 2–4, text-fig. 36.
1986Cordylodus rotundatus Pander; Reference Chen and GongChen and Gong, p. 133, pl. 37, figs. 3, 8, 18, text-fig. 41.
1987Cordylodus angulatus Pander; Reference DongDong, p. 153, pl. 1, figs. 17, 18, 22, text-figs. 3H, J, 4F.
1987Cordylodus rotundatus Pander; Reference DongDong, p. 155, pl. 1, fig. 20 only (not 23), text-fig. 3I only (not K).
1990Cordylodus angulatus Pander; Reference NicollNicoll, figs. 3 (4b–c only), 5 (3a–4c only), 12.
1994Cordylodus angulatus Pander; Reference Ji and BarnesJi and Barnes, p. 31, pl. 5, figs. 1–9.
1994Cordylodus angulatus Pander; Reference Wright and CooperWright and Cooper, p. 471, fig. 17B.
1996Cordylodus angulatus Pander; Reference Lee, Lee and KoLee et al., p. 99, pl. 2, fig. 1.
1998Cordylodus angulatus Pander; Reference Rao and TortelloRao and Tortello, p. 43, pl. 1, figs. 1, 2, 3, 6.
2000Cordylodus angulatus Pander; Reference Zhao, Zhang and XiaoZhao et al., p. 194, pl. 37, figs. 13, 14, 16, 17.
2000Cordylodus rotundatus Pander; Reference Zhao, Zhang and XiaoZhao et al., p. 194, pl. 37, figs. 7–12.
2001Cordylodus angulatus Pander; Reference Pyle and BarnesPyle and Barnes, p. 1397, pl. 1, figs. 3, 4.
2002Cordylodus angulatus Pander; Reference Pyle and BarnesPyle and Barnes, p. 177, pl. 4, figs. 1–3.
2003Cordylodus angulatus Pander; Reference Heinsalu, Kaljo, Kurvits and ViiraHeinsalu et al., p. 147, text-fig. 5, 24 (only).
2004cCordylodus angulatus Pander; Reference Dong, Donoghue, Cheng and LiuDong et al., p. 1201, pl. 2, figs. 17, 28, 30, pl. 4, figs. 4, 6, 12, 14.
2005Cordylodus angulatus Pander; Reference Ortega and AlbanesiOrtega and Albanesi, p. 365, fig. 5 (14).
2007Cordylodus angulatus Pander; Reference Pyle, Barnes and McanallyPyle et al., p. 1735, fig. 11.8, 11.9.
Description.—
Bimembrate apparatus of rounded and compressed elements. Rounded element with cusp and denticles rounded or oval in cross section and composed mostly of white matter. Basal cavity shallow to moderately deep, usually extending not higher than top of posterior denticulate process. Anterior edge of basal cavity strongly concave and tip recurved. Anterobasal margin slightly rounded to well rounded. Posterior process without carina or with a carina on one side. Compressed element symmetrical or subsymmetrical, with long posterior denticulate process. Cusp and denticles compressed laterally with sharp edges. Basal cavity shallow to moderately deep. Anterior edge of basal cavity slightly concave.
Materials.—
12 specimens.
Occurrence.—
Known from the Wa’ergang section, where it occurs in the Cordylodus angulatus Zone (lower part).
Remarks.—
Miller (Reference Miller1980) suggested that the two form-species Cordylodus angulatus and Cordylodus rotundatus belong to two multielement species mainly based on the minor differences between them. However, many conodont workers considered the two form-species to be a single apparatus, because the two form-species commonly occur together. Based on the common association of the two form-species recovered from Hunan, we follow the opinion of the majority.
Cordylodus caseyi Druce and Jones, Reference Druce and Jones1971
Figures 9.21, 9.25, 9.26, 11.4

Figure 11 (1–3, 5–27) Cordylodus intermedius Furnish, Reference Furnish1938; (1) Panjiazui Formation, Wa’ergang section, GMPKU2452, lateral view; (2) Panjiazui Formation, Wa’ergang section, GMPKU2453, lateral view; (3) Panjiazui Formation, Wa’ergang section, GMPKU2454, lateral view; (5) Panjiazui Formation, Wa’ergang section, GMPKU2455, lateral view; (6) Panjiazui Formation, Wa’ergang section, GMPKU2456, lateral view; (7) Panjiazui Formation, Wa’ergang section, GMPKU2457, lateral view; (8) Panjiazui Formation, Wa’ergang section, GMPKU2458, lateral view; (9) Panjiazui Formation, Wa’ergang section, GMPKU2459, lateral view; (10) Panjiazui Formation, Wa’ergang section, GMPKU2460, lateral view; (11) Panjiazui Formation, Wa’ergang section, GMPKU2461, lateral view; (12) Panjiazui Formation, Wa’ergang section, GMPKU2462, lateral view; (13) Panjiazui Formation, Wa’ergang section, GMPKU2463, lateral view; (14) Panjiazui Formation, Wa’ergang section, GMPKU2464, lateral view; (15) Panjiazui Formation, Wa’ergang section, GMPKU2465, lateral view; (16) Panjiazui Formation, Wa’ergang section, GMPKU2466, lateral view; (17) Panjiazui Formation, Wa’ergang section, GMPKU2467, lateral view; (18) Panjiazui Formation, Wa’ergang section, GMPKU2468, lateral view; (19) Panjiazui Formation, Wa’ergang section, GMPKU2469, lateral view; (20) Panjiazui Formation, Wa’ergang section, GMPKU2470, lateral view; (21) Panjiazui Formation, Wa’ergang section, GMPKU2471, lateral view; (22) Panjiazui Formation, Wa’ergang section, GMPKU2472, lateral view; (23) Panjiazui Formation, Wa’ergang section, GMPKU2473, lateral view; (24) Panjiazui Formation, Wa’ergang section, GMPKU2474, lateral view; (25) Panjiazui Formation, Wa’ergang section, GMPKU2475, lateral view; (26) Panjiazui Formation, Wa’ergang section, GMPKU2476, lateral view; (27) Panjiazui Formation, Wa’ergang section, GMPKU2477, lateral view; (4), Cordylodus caseyi Druce and Jones, Reference Druce and Jones1971; Panjiazui Formation, Wa’ergang section, GMPKU2451, lateral view. Relative scale bar represents 180 μm (1–21, 24–27), 145 μm (22), 154 μm (23).
1971Cordylodus caseyi Reference Druce and JonesDruce and Jones, p. 67, pl. 2, figs. 9–12, text-figs. 23d, e.
1990Cordylodus caseyi Druce and Jones; Reference NicollNicoll, p. 543, figs. 3 (3b), 13 (4a–5c), 14 (1–2) only.
1993Cordylodus caseyi Druce and Jones; Reference LandingLanding, p. 14, fig. 8, 3–13, p. 15, fig. 9, 1–11.
2001Cordylodus caseyi Druce and Jones; Reference Pyle and BarnesPyle and Barnes, p. 1397, pl. 1, fig. 6.
2002Cordylodus caseyi Druce and Jones; Reference Pyle and BarnesPyle and Barnes, p. 177, pl. 4, figs. 4–7.
2003Cordylodus caseyi Druce and Jones; Reference Landing, Westrop and van Aller HernickLanding et al., p. 86, fig. 7, 16 (only).
Description.—
The element is asymmetrical. The long cusp is reclined, oval in cross-section. The base is greatly expanded to give a triangular cross section in the basal view. The basal cavity is moderately large with anterior edge concave or straight. A conspicuous keel is developed on the lateral margin of the cusp and base.
Materials.—
26 specimens.
Occurrence.—
Known from the Wa’ergang section, where it occurs in the Cordylodus lindstromi Zone.
Remarks.—
This species is characterized by its expanded base with a triangular cross-section and the keel developed on the lateral margin of the cusp and base. Only rounded elements have been found for this species.
Cordylodus intermedius Furnish, Reference Furnish1938
Figures 10.3–10.6, 11.1–11.3, 11.5–11.27, 12.1–12.4, 12.7

Figure 12 (1–4, 7) Cordylodus intermedius Furnish, Reference Furnish1938; (1) Panjiazui Formation, Wa’ergang section, GMPKU2478, lateral view; (2) Panjiazui Formation, Wa’ergang section, GMPKU2479, lateral view; (3) Panjiazui Formation, Wa’ergang section, GMPKU2480, lateral view; (4) Panjiazui Formation, Wa’ergang section, GMPKU2481, lateral view; (7) Panjiazui Formation, Wa’ergang section, GMPKU2482, lateral view; (5, 6, 8–26) Cordylodus lindstromi Druce and Jones, Reference Druce and Jones1971; (5) Panjiazui Formation, Wa’ergang section, GMPKU2483, lateral view; (6) Panjiazui Formation, Wa’ergang section, GMPKU2484, lateral view; (8) Panjiazui Formation, Wa’ergang section, GMPKU2485, lateral view; (9) Panjiazui Formation, Wa’ergang section, GMPKU2486, lateral view; (10) Panjiazui Formation, Wa’ergang section, GMPKU2487, lateral view; (11) Panjiazui Formation, Wa’ergang section, GMPKU2488, lateral view; (12) Panjiazui Formation, Wa’ergang section, GMPKU2489, lateral view; (13) Panjiazui Formation, Wa’ergang section, GMPKU2490, lateral view; (14) Panjiazui Formation, Wa’ergang section, GMPKU2491, lateral view; (15) Panjiazui Formation, Wa’ergang section, GMPKU2492, lateral view; (16) Panjiazui Formation, Wa’ergang section, GMPKU2493, lateral view; (17) Panjiazui Formation, Wa’ergang section, GMPKU2494, lateral view; (18) Panjiazui Formation, Wa’ergang section, GMPKU2496, lateral view; (19) Panjiazui Formation, Wa’ergang section, GMPKU2497, lateral view; (20) Panjiazui Formation, Wa’ergang section, GMPKU2498, lateral view; (21) Panjiazui Formation, Wa’ergang section, GMPKU2499, lateral view; (22) Panjiazui Formation, Wa’ergang section, GMPKU2500, lateral view; (23) Panjiazui Formation, Wa’ergang section, GMPKU2501, lateral view; (24) Panjiazui Formation, Wa’ergang section, GMPKU2502, lateral view; (25) Panjiazui Formation, Wa’ergang section, GMPKU2503, lateral view; (26) Panjiazui Formation, Wa’ergang section, GMPKU2504, lateral view. Relative scale bar represents 128 μm (1–7, 11–20, 22, 24–26), 114 μm (8–10, 21, 23).
1938Cordylodus intermedius Reference FurnishFurnish, p. 338, pl. 42, fig. 31, text-fig. 2c.
1955Cordylodus prion Reference LindströmLindström, p. 552, pl. 5, figs. 14–16.
1971Cordylodus intermedius Furnish; Reference Druce and JonesDruce and Jones, p. 68, pl. 3, figs. 1–3, text-figs. 23f, g.
1971Cordylodus oklahomensis Müller; Reference Druce and JonesDruce and Jones, p. 69, pl. 5, figs. 6–7, text-fig. 23j.
1971Cordylodus prion Lindström; Reference LindströmLindström, pl. 2, figs. 1–7, text-figs. 23i, k–o.
1971Cordylodus proavus Müller; Reference Druce and JonesDruce and Jones, p. 70, text-fig. 23p, pl. 1, fig. 1 only (not figs. 2–6). (part)
1973Cordylodus intermedius Furnish; Reference MüllerMüller, p. 30, pl. 10, figs. 1–3, text-figs. 2c, 4a, b.
1973Cordylodus prion Lindström; Reference MüllerMüller, p. 33, text-figs. 2E, 8, pl. 10, fig. 4.
1974Cordylodus prion Lindström; Reference van Wamelvan Wamel, p. 59, pl. 1, figs. 8–9.
1980Cordylodus intermedius Furnish; Reference MillerMiller, p. 17, pl. 1, fig. 16, text-fig. 4L.
1980Cordylodus intermedius Furnish; Reference Landing, Ludvigen and von BitterLanding et al., p. 19, figs. 5E, 6A, B.
1982Cordylodus intermedius Furnish; s.f. Reference RepetskiRepetski, p. 17, pl. 5, fig. 2, text-fig. 4 (M).
1983Cordylodus intermedius Furnish; Reference An, Zhang, Xiang, Zhang, Xu, Zhang, Jiang, Yang, Lin, Cui and YangAn et al., p. 85, pl. 7, fig. 16.
1985Cordylodus intermedius Furnish; Reference WangWang, p. 215, pl. 1, figs. 12, 13, 15, 14 (?), pl. 7, figs. 22–24, pl. 9, figs. 5–7, pl. l l, figs. 5–7; pl. 13, figs. 22, 23 (not pl. 14, fig. 4). (part)
1985Cordylodus intermedius Furnish; Reference DongDong, p. 394, pl. 2, fig. 6, pl. 3, fig. 10, text-fig. 1k.
1986Cordylodus intermedius Furnish; Reference Chen and GongChen and Gong, p. 127, pl. 35, figs. 2, 3, 6, 9, pl. 36, fig. 7, pl. 37, figs. 1, 2, 4–7, 10–11, 13, 15–17, pl. 38, figs. 2, 3, 7–8, 13, 15, text-fig. 38.
1987Cordylodus intermedius Furnish; Reference DongDong, p. 154, pl. 1, figs. 14, 15, 19, 25, text-figs. 3E, F, L, 4C.
1987Cordylodus intermedius Furnish; Bagnoli et al., p. 153, pl. 1, figs. 15–18.
1994Cordylodus intermedius Furnish; Reference Ji and BarnesJi and Barnes, p. 32, pl. 5, figs. 10–18.
1998Cordylodus intermedius Furnish; Reference Rao and TortelloRao and Tortello, p. 43, pl. 1, fig. 7.
2000Cordylodus intermedius Furnish; Reference Zhao, Zhang and XiaoZhao et al., p. 195, pl. 37, figs. 15, 21.
2000“Cordylodus” intermedius Furnish; Reference Pyle and BarnesDubinina, p. 188, pl. 9, fig. 25.
? 2000“Cordylodus” lenzi Müller; Reference Pyle and BarnesDubinina, p. 188, pl. 9, fig. 21.
2002Cordylodus intermedius Furnish; Reference Pyle and BarnesPyle and Barnes, p. 47, pl. 4, figs. 15–17.
2004cCordylodus intermedius Furnish; Reference Dong, Donoghue, Cheng and LiuDong et al., pl. 2, figs. 8, 9, 19, 21, 24, pl. 4, figs. 5, 13.
Description.—
Bimembrate apparatus of rounded and compressed elements. Rounded element symmetrical or subsymmetrical, possessing a distinctive basal cavity with anterior edge concave or straight. Compressed element may be symmetrical or asymmetrical due to lateral bending of cusp and presence of the slight carina on inner side of basal margin. Cusp and denticles laterally compressed and sharp edges, denticles fused at base.
Materials.—
260 specimens.
Occurrence.—
Known from the Wa’ergang section, where it ranges from the Cordylodus intermedius Zone through the Cordylodus lindstromi Zone (upper part).
Remarks.—
The overwhelming majority of conodont workers believed that Cordylodus intermedius is the evolutionary intermediate between Cordylodus proavus and Cordylodus angulatus. Nevertheless, Nicoll (Reference Nicoll1990) proposed that C. intermedius is an invalid species, and that much of the material previously assigned to C. intermedius should reassigned to C. angulatus and that some of the stratigraphically older specimens probably should be assigned to C. proavus. He also replaced the C. intermedius Zone by a lower Hirsutodontus simplex Zone and an upper Clavohamamulus hintzei Zone. However, in terms of stratigraphical range, the FAD of C. intermedius is just between that of C. proavus and that of C. angulatus. In terms of morphology, the anterior edge of basal cavity is concave or straight in C. intermedius, whereas it is convex in C. proavus. Therefore, the reassignment of the stratigraphically older specimens of C. intermedius to C. proavus is absolutely inconsistent with the orginal definition of the two form-species, even if the reassignment of some specimens of C. intermedius to C. angulatus sounds plausible. So far as it goes, Nicoll’s (Reference Nicoll1990) proposal has never been accepted, and C. intermedius is unequivocally a valid species, while C. intermedius Zone is valid and useful for the work on Cambrian–Ordovician boundary (Cooper et al., Reference Cooper, Nowlan and Williams2001; Terfelt et al., Reference Terfelt, Bagnoli and Stouge2012)
Cordylodus lindstromi Druce and Jones, Reference Druce and Jones1971
Figures 12.5, 12.6, 12.8–12.26, 13.1–13.15

Figure 13 Conodont images using oil immersion techniques (Dong et al., 2005c) with Differential Interference Contrast (Nomarski) illumination, showing the shapes of the biapical basal cavities. (1–15) Cordylodus lindstromi Druce and Jones, Reference Druce and Jones1971; (1) the same specimen as Figure 12.10; (2) the same specimen as Figure 12.8; (3) the same specimen as Figure 12.11; (4), the same specimen as Figure 12.13; (5) the same specimen as Figure 12.12; (6) the same specimen as Figure 12.14; (7) the same specimen as Figure 12.23; (8) the same specimen as Figure 12.15; (9) the same specimen as Figure 12.16; (10) the same specimen as Figure 12.17; (11) the same specimen as Figure 12.19; (12) the same specimen as Figure 12.18: (13) the same specimen as Figure 12.20; (14) the same specimen as Figure 12.24; (15) the same specimen as Figure 12.25. Relative scale bar represents 80 μm (1, 5–7, 11, 13), 67 μm (2), 121 μm (3, 9, 10, 12, 15), 92 μm (4), 58 μm (8), 103 μm (14).
1971Cordylodus lindstromi Reference Druce and JonesDruce and Jones, p. 68, pl. 1, figs. 7–9, text-fig. 23h, pl. 2, fig. 8.
1973Cordylodus lindstromi Druce and Jones; Reference MüllerMüller, p. 22, text-figs. 2d, 6a, b, pl. 9, figs. 10, 11.
1982Cordylodus lindstromi Druce and Jones; s.f. Reference RepetskiRepetski, p. 17, pl. 5, figs. 4–5, text-fig. 4 (O).
1983Cordylodus lindstromi Druce and Jones; Reference An, Zhang, Xiang, Zhang, Xu, Zhang, Jiang, Yang, Lin, Cui and YangAn et al., p. 86, pl. 7, figs. 17–19.
1985Cordylodus lindstromi Druce and Jones; Reference WangWang, p. 216, pl. 1, fig. 6, pl. 13, fig. 24.
1986Cordylodus lindstromi Druce and Jones; Reference Chen and GongChen and Gong, p. 129, pl. 34, figs. 1, 5–8, text-fig. 39.
1987Cordylodus lindstromi Druce and Jones; Reference DongDong, p. 154, pl. 1, fig. 30, text-fig. 3, G.
1990Cordylodus lindstromi Druce and Jones; Reference NicollNicoll, p. 545, fig. 3, (2b only), fig. 16, (7–14 only), fig. 17 (not fig. 18).
1991Cordylodus lindstromi Druce and Jones; Reference NicollNicoll, p. 231, figs. 2.3, 2.4, fig. 4. (part)
1994Cordylodus lindstromi Druce and Jones; Reference Ji and BarnesJi and Barnes, p. 32, pl. 5, figs. 19–22.
? 1999Cordylodus lindstromi Druce and Jones; Reference Parsons and ClarkParsons and Clark, p. 7, figs. 26–27.
2000Cordylodus lindstromi Druce and Jones; Reference Göncüoğlu and KozurGöncüoğlu and Kozur, p. 599, fig. 4, g, p. 601, fig. 5, a, g.
2000“Cordylodus” lindstromi Druce and Jones; Reference Pyle and BarnesDubinina, p. 188, pl. 9, figs. 15, ?16–17.
2001Cordylodus lindstromi Druce and Jones; Reference Pyle and BarnesPyle and Barnes, p. 1397, pl. 1, fig. 7.
2002Cordylodus lindstromi Druce and Jones; Reference Pyle and BarnesPyle and Barnes, p. 48, pl. 4, figs. 18–21.
2003Cordylodus lindstromi Druce and Jones; Reference Heinsalu, Kaljo, Kurvits and ViiraHeinsalu et al., p. 147, text-fig. 5, 13–14 (only, not 15–20). (part)
2004cCordylodus lindstromi Druce and Jones; Reference Dong, Donoghue, Cheng and LiuDong et al., pl. 2, figs. 14, 16, 22, 23, pl. 4, figs. 7, 15.
2007Cordylodus lindstromi Druce and Jones; Reference Pyle, Barnes and McanallyPyle et al., p. 1733, fig. 10, 3–4.
Description.—
Bimembrate apparatus of rounded and compressed elements. Rounded elements and compressed elements symmetrical, with distinctive basal cavities. Secondary apex of basal cavity extends into the first, and rarely the second, denticle. The anterior edge of basal cavity is convex.
Materials.—
200 specimens.
Occurrence.—
Known from the Wa’ergang section, where it where it occurs in the Cordylodus lindstromi Zone.
Remarks.—
The specimens at hand agree well with Cordylodus lindstromi.
Cordylodus proavus Müller, Reference Müller1959
Figures 9.16, 14.1–14.23
1959Cordylodus proavus Reference MüllerMüller, p. 448, pl. 15, figs. 11, 12, 18, text-fig. 3B.
1959Cordylodus oklahomensis Reference MüllerMüller, p. 447, text-fig. 3A, pl. 15, figs. 15, 16.
1969Cordylodus proavus Müller; Reference MillerMiller, p. 424, pl. 65, figs. 37–45, text-fig. 3D.
1969Cordylodus oklahomensis Müller; Reference MillerMiller, p. 423, text-fig. 3, I, pl. 65, figs. 46–53.
1971Cordylodus proavus Müller; Reference Druce and JonesDruce and Jones, p. 70, pl. 1, figs. 2–6 (not 1), text-fig. 23q, r (not p). (part)
1971Cordylodus cf. C. proavus Müller; Reference Druce and JonesDruce and Jones, p. 71, pl. 1, figs. 10, 11 (?), 12, text-fig. 23s.
1971Cordylodus proavus Müller; Reference Ethington and ClarkEthington and Clark, pl. 1, fig. 19.
1971Cordylodus oklahomensis Müller; Reference Ethington and ClarkEthington and Clark, pl. 1, fig. 24.
1973Cordylodus proavus Müller; Reference MüllerMüller, p. 35, text-figs. 2A, 9, pl. 9, figs. 1–9.
1973Cordylodus oklahomensis Müller; Reference MüllerMüller, p. 33, text-figs. 2b, 7a, b, pl. 9, figs. 12, 13.
1974Cordylodus angulatus Pander; Reference van Wamelvan Wamel, pl. 1, fig. 5 (not figs. 6, 7). (part)
1980Cordylodus proavus Müller; Reference MillerMiller, p. 19–20, text-figs. 4G, H, pl. 1, figs. 14, 15.
1983Cordylodus proavus Müller; Reference An, Zhang, Xiang, Zhang, Xu, Zhang, Jiang, Yang, Lin, Cui and YangAn et al., p. 87–88, pl. 7, figs. l–5, 8, 11.
1983Cordylodus aff. C. proavus Müller; Reference An, Zhang, Xiang, Zhang, Xu, Zhang, Jiang, Yang, Lin, Cui and YangAn et al., p. 88, pl. 7, figs. 6, 7, 9, 10, text-fig. 10 (3).
1985Cordylodus proavus Müller; Reference DongDong, p. 395–396, pl. 1, figs. 1, 2, text-figs. 1 (A, B).
1985Cordylodus proavus Müller; Reference WangWang, p. 217, pl. 1, figs. 10, 18–23, pl. 7, figs. 15–20, pl. 9, figs. 9–12, pl. 11, figs. 2–4, 19–21, figs. 2–4, 19–21, pl. 13, figs. 26, 31, pl. 14, figs. 11–16.
1986Cordylodus proavus Müller; Reference Chen and GongChen and Gong, p. 130, pl. 35, figs. 1, 4–5, 7–8, 10–14, pl. 36, figs. 1–6, 8–24, pl. 37, figs. 9, 12, 14, pl. 38, figs. 1, 4, 6, 9, 11–12, 14, 16–18, text-fig. 40.
1987Cordylodus proavus Müller; Reference DongDong, p. 155, pl. 1, figs. 7–10, text-fig. 3D.
1990Cordylodus proavus Müller; Reference NicollNicoll, p. 550, figs. 3 (1b–d only, not 1a), 19 (2–4 only, not 1), 20–22. (part)
1993Cordylodus proavus Müller; Reference LandingLanding, p. 14, figs. 8, 16, figs. 9, 14.
1996Cordylodus proavus Müller; Reference Lee, Lee and KoLee et al., p. 97, pl. 1, figs. 4–5, 9–10.
? 1999Cordylodus proavus Müller; Reference Parsons and ClarkParsons and Clark, p. 7, fig. 5 (24–25).
2000Cordylodus proavus Müller; Reference Pyle and BarnesDubinina, p. 190, pl. 9, figs. 1–4, 6, 8–10, 14, 19, 20.
2000Cordylodus proavus Müller; Reference Zhao, Zhang and XiaoZhao et al., p. 195, pl. 37, figs. 18–20.
2001Cordylodus proavus Müller; Reference LeeLee, p. 451, fig. 7 (20–23).
2002aCordylodus proavus Müller; Reference LeeLee, p. 165, pl. 2, figs. 31, 32.
2002bCordylodus proavus Müller; Reference LeeLee, p. 27, pl. 2, figs. 5, 6.
2002Cordylodus proavus Müller; Reference Pyle and BarnesPyle and Barnes, p. 49, pl. 5, figs. 4–6.
2003Cordylodus proavus Müller; Reference Landing, Westrop and van Aller HernickLanding et al., p. 86, figs. 7, 11–13.
2003Cordylodus proavus Müller; Reference Heinsalu, Kaljo, Kurvits and ViiraHeinsalu et al., p. 147, text-fig. 5 (1–4).
2004Cordylodus proavus Müller; Reference LeeLee, p. 60, fig. 5, BB–CC.
2004cCordylodus proavus Müller; Reference Dong, Donoghue, Cheng and LiuDong et al., pl. 2, figs. 11, 12, 15, 20, 29, pl. 4, figs. 3, 11.
2006Cordylodus proavus Müller; Reference Miller, Ethington, Evans, Holmer, Loch, Popov, Repetski, Ripperdan and TaylorMiller et al., p. 394, fig. 9 A, B.
2007Cordylodus proavus Müller; Reference Landing, Westrop and KeppieLanding et al., p. 917, fig. 5 (h–l).
2007Cordylodus proavus Müller; Reference Pyle, Barnes and McanallyPyle et al., p. 1733, fig. 10 (1–2).
Description.—
Bimembrate apparatus of rounded and compressed elements, possessing deep basal cavities, the anterior edge of basal cavities is convex, with the tips of basal cavities extending higher than the base of most posterior denticles up to near the apex of the element. Rounded element symmetrical or subsymmetrical. Cusp and denticles round or oval in cross section, composed of white matter. Denticles separated from one another at base. Compressed element asymmetrical due to lateral bending of cusp and due to a carina near the base of inner side. Cusp and denticles composed of white matter, compressed laterally, with keeled anterior and posterior edges. Denticles fused at bases.
Materials.—
600 specimens.
Occurrence.—
Known from the Wa’ergang section, where it ranges from the Cordylodus proavus Zone through the Cordylodus lindstromi Zone (upper part).
Cordylodus prolindstromi Nicoll, Reference Nicoll1991
Figures 14.24–14.30, 15.1–15.35, 16.1–16.26

Figure 14 (1–23) Cordylodus proavus Müller, Reference Müller1959; (1) Shenjiawan Formation, Wa’ergang section, GMPKU2505, lateral view; (2) Shenjiawan Formation, Wa’ergang section, GMPKU2506, lateral view; (3) Shenjiawan Formation, Wa’ergang section, GMPKU2507, lateral view; (4) Panjiazui Formation, Wa’ergang section, GMPKU2508, lateral view; (5) Panjiazui Formation, Wa’ergang section, GMPKU2509, lateral view; (6) Panjiazui Formation, Wa’ergang section, GMPKU2510, lateral view; (7) Panjiazui Formation, Wa’ergang section, GMPKU2511, lateral view; (8) Panjiazui Formation, Wa’ergang section, GMPKU2512, lateral view; (9) Panjiazui Formation, Wa’ergang section, GMPKU2513, lateral view; (10) Panjiazui Formation, Wa’ergang section, GMPKU2514, lateral view; (11) Panjiazui Formation, Wa’ergang section, GMPKU2515, lateral view; (12) Panjiazui Formation, Wa’ergang section, GMPKU2516, lateral view; (13) Panjiazui Formation, Wa’ergang section, GMPKU2517, lateral view; (14) Panjiazui Formation, Wa’ergang section, GMPKU2518, lateral view; (15) Panjiazui Formation, Wa’ergang section, GMPKU2519, lateral view; (16) Panjiazui Formation, Wa’ergang section, GMPKU2520, lateral view; (17) Panjiazui Formation, Wa’ergang section, GMPKU2521, lateral view; (18) Panjiazui Formation, Wa’ergang section, GMPKU2522, lateral view; (19) Panjiazui Formation, Wa’ergang section, GMPKU2523, lateral view; (20) Panjiazui Formation, Wa’ergang section, GMPKU2524, lateral view; (21) Panjiazui Formation, Wa’ergang section, GMPKU2525, lateral view; (22) Panjiazui Formation, Wa’ergang section, GMPKU2526, lateral view; (23) Panjiazui Formation, Wa’ergang section, GMPKU2527, lateral view; (24–30) Cordylodus prolindstromi Nicoll, Reference Nicoll1991; (24) Panjiazui Formation, Wa’ergang section, GMPKU2528, lateral view; (25) Panjiazui Formation, Wa’ergang section, GMPKU2529, lateral view; (26) Panjiazui Formation, Wa’ergang section, GMPKU2530, lateral view; (27) Panjiazui Formation, Wa’ergang section, GMPKU2531, lateral view; (28) Panjiazui Formation, Wa’ergang section, GMPKU2532, lateral view; (29) Panjiazui Formation, Wa’ergang section, GMPKU2533, lateral view; (30) Panjiazui Formation, Wa’ergang section, GMPKU2534, lateral view. Relative scale bar represents 145 μm (1, 3–10, 12–16, 19, 21, 22, 24, 27, 29), 136 μm (2, 11, 17, 18, 20, 23, 25, 26, 28, 30).

Figure 15 (1–35) Cordylodus prolindstromi Nicoll, Reference Nicoll1991; (1) Panjiazui Formation, Wa’ergang section, GMPKU2535, lateral view; (2) Panjiazui Formation, Wa’ergang section, GMPKU2536, lateral view; (3) Panjiazui Formation, Wa’ergang section, GMPKU2537, lateral view; (4) Panjiazui Formation, Wa’ergang section, GMPKU2538, lateral view; (5) Panjiazui Formation, Wa’ergang section, GMPKU2539, lateral view; (6) Panjiazui Formation, Wa’ergang section, GMPKU2540, lateral view; (7) Panjiazui Formation, Wa’ergang section, GMPKU2541, lateral view; (8) Panjiazui Formation, Wa’ergang section, GMPKU2542, lateral view; (9) Panjiazui Formation, Wa’ergang section, GMPKU2543, lateral view; (10) Panjiazui Formation, Wa’ergang section, GMPKU2544, lateral view; (11) Panjiazui Formation, Wa’ergang section, GMPKU2545, lateral view; (12) Panjiazui Formation, Wa’ergang section, GMPKU2546, lateral view; (13) Panjiazui Formation, Wa’ergang section, GMPKU2547, lateral view; (14) Panjiazui Formation, Wa’ergang section, GMPKU2548, lateral view; (15) Panjiazui Formation, Wa’ergang section, GMPKU2549, lateral view; (16) Panjiazui Formation, Wa’ergang section, GMPKU2550, lateral view; (17) Panjiazui Formation, Wa’ergang section, GMPKU2551, lateral view; (18) Panjiazui Formation, Wa’ergang section, GMPKU2552, lateral view; (19) Panjiazui Formation, Wa’ergang section, GMPKU2553, lateral view; (20) Panjiazui Formation, Wa’ergang section, GMPKU2554, lateral view; (21) Panjiazui Formation, Wa’ergang section, GMPKU2555, lateral view; (22) Panjiazui Formation, Wa’ergang section, GMPKU2556, lateral view; (23) Panjiazui Formation, Wa’ergang section, GMPKU2557, lateral view; (24) Panjiazui Formation, Wa’ergang section, GMPKU2558, lateral view; (25) Panjiazui Formation, Wa’ergang section, GMPKU2559, lateral view; (26) Panjiazui Formation, Wa’ergang section, GMPKU2560, lateral view; (27) Panjiazui Formation, Wa’ergang section, GMPKU2561, lateral view; (28) Panjiazui Formation, Wa’ergang section, GMPKU2562, lateral view; (29) Panjiazui Formation, Wa’ergang section, GMPKU2563, lateral view; (30) Panjiazui Formation, Wa’ergang section, GMPKU2564, lateral view; (31) Panjiazui Formation, Wa’ergang section, GMPKU2565, lateral view; (32) Panjiazui Formation, Wa’ergang section, GMPKU2566, lateral view; (33) Panjiazui Formation, Wa’ergang section, GMPKU2567, lateral view; (34) Panjiazui Formation, Wa’ergang section, GMPKU2568, lateral view; (35) Panjiazui Formation, Wa’ergang section, GMPKU2569, lateral view; Relative scale bar represents 150 μm (1–3, 5–23, 25–27, 30–35), 137 μm (4, 24, 28), 120 μm (29).

Figure 16 Conodont images using oil immersion techniques (Dong et al., 2005c) with Differential Interference Contrast (Nomarski) illumination, showing the shapes of the biapical basal cavities. (1–26) Cordylodus prolindstromi Nicoll, Reference Nicoll1991; (1) the same specimen as Figure 15.10; (2) the same specimen as Figure 15.4; (3) the same specimen as Figure 15.5; (4) the same specimen as Figure 15.6; (5) the same specimen as Figure 15.11; (6) the same specimen as Figure 15.13; (7) the same specimen as Figure 15.14; (8) the same specimen as Figure 15.15; (9) the same specimen as Figure 15.16; (10) the same specimen as Figure 15.17; (11) the same specimen as Figure 15.19; (12) the same specimen as Figure 15.20; (13) the same specimen as Figure 15.21; (14) the same specimen as Figure 15.22; (15) the same specimen as Figure 15.23; (16) the same specimen as Figure 15.25; (17) the same specimen as Figure 15.26; (18) the same specimen as Figure 15.27; (19) the same specimen as Figure 15.28; (20) the same specimen as Figure 15.29; (21) the same specimen as Figure 15.30; (22) the same specimen as Figure 15.31; (23) the same specimen as Figure 15.32; (24) the same specimen as Figure 15.33; (25) the same specimen as Figure 15.34; (26) the same specimen as Figure 15.35. Relative scale bar represents 168 μm (1, 4), 149 μm (2, 3, 6, 10, 12, 17, 18, 21, 22, 24), 121 μm (5, 8, 13, 16, 26), 109 μm (7, 14, 19), 134 μm (9, 11, 23, 25), 98 μm (15), 101 μm (20).
1991Cordylodus prolindstromi Reference NicollNicoll, p. 233, figs. 2 (4), 13 (3–4 only, not 1–2), 14–16.
Materials.—
250 specimens.
Occurrence.—
Known from Wa’ergang section where it ranges from the Cordylodus intermedius Zone through the Cordylodus lindstromi Zone.
Remarks.—
Nicoll (Reference Nicoll1991) erected Cordylodus prolindstromi and suggested it is the ancestor of Cordylodus lindstromi. The main difference between them is the shape of the second apex of basal cavity. According to Nicoll (Reference Nicoll1991), in C. lindstromi the secondary apex of the basal cavity is pointed, whereas in C. prolindstromi, it is truncated by a flat top. In many specimens of C. prolindstromi, the tip of the secondary apex of the basal cavity appears to be truncated by a flat top when observed under the ordinary binocular microscope. However, specimens from Hunan have been examined using the oil immersion technique (Dong et al., Reference Dong, Donoghue and Repetski2005c). Photomicrographs of these specimens taken with a differential interference contrast (Nomarski) microscope indicate that the shape of the secondary apex is irregular, i.e, it is neither pointed nor truncated by a flat top (Fig. 15). So, the truncated, flat-top shape of the secondary apex may be a preservational artifact. In light of this, C. prolindstromi may be a junior synonym of C. lindstromi. Nevertheless, because the type specimens of C. prolindstromi from Australia have not been examined using the oil immersion technique, we provisionally retain the name of this species.
Genus Dasytodus Chen and Gong, Reference Chen and Gong1986
Type species.—
Proconodontus transmutatus Xu and Xiang in An et al., Reference An, Zhang, Xiang, Zhang, Xu, Zhang, Jiang, Yang, Lin, Cui and Yang1983.
Dasytodus transmutatus (Xu and Xiang in An et al., Reference An, Zhang, Xiang, Zhang, Xu, Zhang, Jiang, Yang, Lin, Cui and Yang1983)
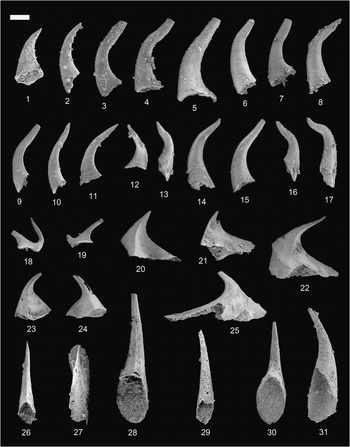
Figure 17 (1, 2) Dasytodus transmutatus (Xu and Xiang, 1983); (1) Shenjiawan Formation, Wangcun section, GMPKU2570, lateral view; (2) Shenjiawan Formation, Wa’ergang section, GMPKU2571, lateral view; (3–17) Eoconodontus notchpeakensis Miller, Reference Miller1980; (3) Shenjiawan Formation, Wangcun section, GMPKU2572, lateral view; (4) Shenjiawan Formation, Wangcun section, GMPKU2573, lateral view; (5) Shenjiawan Formation, Wa’ergang section, GMPKU2574, lateral view; (6) Shenjiawan Formation, Wa’ergang section, GMPKU2575, lateral view; (7) Shenjiawan Formation, Wa’ergang section, GMPKU2576, lateral view; (8) Shenjiawan Formation, Wa’ergang section, GMPKU2577, lateral view; (9) Shenjiawan Formation, Wa’ergang section, GMPKU2578, lateral view; (10) Shenjiawan Formation, Wa’ergang section, GMPKU2579, lateral view; (11) Shenjiawan Formation, Wa’ergang section, GMPKU2580, lateral view; (12) Shenjiawan Formation, Wa’ergang section, GMPKU2581, lateral view; (13) Shenjiawan Formation, Wa’ergang section, GMPKU2582, lateral view; (14) Shenjiawan Formation, Wa’ergang section, GMPKU2583, lateral view; (15) Shenjiawan Formation, Wa’ergang section, GMPKU2584, lateral view; (16) Shenjiawan Formation, Wa’ergang section, GMPKU2585, lateral view; (17) Shenjiawan Formation, Wa’ergang section, GMPKU2586, lateral view; (18, 19) Eodentatus bicuspatus Nicoll and Shergold, Reference Nicoll and Shergold1991; (18) Shenjiawan Formation, Wa’ergang section, GMPKU2587, ramiform element, lateral view; (19) Shenjiawan Formation, Wa’ergang section, GMPKU2588, ramiform element, lateral view; (20–25) Furnishina bigeminata Dong, Reference Dong1993; (20) Huaqiao Formation, Wangcun section, GMPKU2589, outer lateral view; (21) Huaqiao Formation, Wangcun section, GMPKU2590, inner lateral view; (22) Huaqiao Formation, Wangcun section, GMPKU2591, outer lateral view; (23) Chefu Formation, Wangcun section, GMPKU2592, outer lateral view; (24) Chefu Formation, Wangcun section, GMPKU2593, outer lateral view; (25) Bitiao Formation, Wangcun section, GMPKU2594, lateral view; (26–31) Furnishina dayangchaensis Chen and Gong, Reference Chen and Gong1986; (26) Bitiao Formation, Wangcun section, GMPKU2595, posterior view; (27) Bitiao Formation, Wangcun section, GMPKU2596, cluster; (28) Bitiao Formation, Wangcun section, GMPKU2597, posterior view; (29) Bitiao Formation, Wangcun section, GMPKU2598, oblique posterior view; (30) Shenjiawan Formation, Wa’ergang section, GMPKU2599, posterior view; (31) Shenjiawan Formation, Wa’ergang section, GMPKU2600, oblique posterior view. Relative scale bar represents 126 μm (1, 23), 135 μm (2–22, 24–31).
1983Proconodontus transmutatus Xu and Xiang in Reference An, Zhang, Xiang, Zhang, Xu, Zhang, Jiang, Yang, Lin, Cui and YangAn et al., p. 128, pl. 3, figs. 14–16, pl. 33, fig. 3, text-fig. 9 (22).
1986Dasytodus transmutatus (Xu and Xiang); Reference Chen and GongChen and Gong, p. 135, pl. 28, figs. 1, 3–8, 11, text-fig. 43.
2000Hirsutodontus transmutatus (Xu and Xiang); Duninina, p. 187, pl. 6, figs. 3, 3a, not 6. (part)
2001Teridontus transmutatus (Xu and Xiang); Reference LeeLee, p. 449, fig. 6.27, 6.28.
2002aDasytodus transmutatus (Xu and Xiang); Reference LeeLee, p. 165, pl. 2, fig. 17.
2002bTeridontus transmutatus (Xu and Xiang); Reference LeeLee, p. 27, pl. 2, fig. 2.3.
2008aDasytodus transmutatus (Xu and Xiang); Reference LeeLee, p. 228, fig. 1 (6).
Description.—
Coniform elements, proclined and asymmetrical. Cusp short with rounded cross section. Base long and flaring downwards. Small spines scattered mainly on the base but also on the cusp. Basal cavity deep, with the apex extending up to slightly below the point of flexure of the cusp.
Comparison.—
Dasytodus transmutatus differs from Granatodontus ani mainly in the spiny surface ornamentation in the former rather than tiny nodular surface ornamentation in the latter.
Materials.—
Six specimens.
Occurrence.—
Known from the Wangcun and Wa’ergang sections, where it ranges from the Proconodontus Zone through the Cordylodus proavus Zone.
Remarks.—
The specimens at hand agree closely with those illustrated and described by Chen and Gong (Reference Chen and Gong1986), except that spines are scattered also on the cusp in the Hunan material. This difference is considered here to represent intraspecific variation.
Genus Eoconodontus Miller, Reference Miller1980
Type species.—
Proconodontus notchpeakensis Miller, Reference Miller1969.
Eoconodontus notchpeakensis (Miller, Reference Miller1969)
1967Oneotodus nakamurai Reference NogamiNogami, p. 216, text-fig. 3D, E (not text-fig. 3A–C), pl. 1, figs. 10, 11 (not figs. 9, 12, 13). (part)
1969Proconodontus notchpeakensis Reference MillerMiller, p. 438, text-fig. 5G, pl. 66, figs. 21–29.
1969Proconodontus carinatus Reference MillerMiller, p. 437, text-fig. 5I, pl. 66, figs. 15–20 (not figs. 13, 14). (part)
1971Oneotodus nakamurai Nogami; Reference Druce and JonesDruce and Jones, p. 82, text-fig. 26j, pl. 10, figs. 1, 2, 5, 6 (not figs. 3, 4, 7, 8). (part)
1973Proconodontus notchpeakensis Miller; Reference MüllerMüller, p. 43, pl. 4, fig. 6.
1975Oneotodus nakamurai Nogami; Reference LeeLee, p. 81, text-figs. 2E, G, pl. 1, figs. 6, 9, 10.
1977Oneotodus nakamurai Nogami; Reference Abaimova and MarkovAbaimova and Markov, p. 92, pl. 14, figs. 12–14, 16.
1980Eoconodontus notchpeakensis (Miller); Reference MillerMiller, p. 22, pl. 1, figs. 10–12, text-figs. 3D, E.
1982Proconodontus notchpeakensis Miller; Reference AnAn, p. 142, pl. 8, fig. 2, pl. 13, figs. 1–11.
1983Proconodontus notchpeakensis Miller; Reference An, Zhang, Xiang, Zhang, Xu, Zhang, Jiang, Yang, Lin, Cui and YangAn et al., p. 127, pl. 5, figs. 10–12 (not figs. 19, 20). (part)
1985Eoconodontus notchpeakensis (Miller); Reference WangWang, p. 223, pl. 3, figs. 8–10, pl. 4, figs. 10, 11, pl. 7, figs. 1–3, pl. 13, figs. 4, 13, 14, pl. 14, fig. 18.
1985Eoconodontus notchpeakensis (Miller); Reference DongDong, p. 399, pl. 1, figs. 3, 4, text-figs. 1C–D.
1986Eoconodontus notchpeakensis (Miller); Reference Chen and GongChen and Gong, p. 140, pl. 19, fig. 13, pl. 31, figs. 2, 6, 8, pl. 32, figs. 2, 4, 6–7, 9, 11, pl. 33, figs. 2, 8, text-fig. 48.
1987Eoconodontus notchpeakensis (Miller); Reference DongDong, p. 163, pl. 1, figs. 1, 2, pl. 3, figs. 20, 21.
1993Eoconodontus notchpeakensis (Miller); Reference LandingLanding, p. 6, fig. 4 (4, 6, 9).
1996Eoconodontus notchpeakensis (Miller); Reference Lee, Lee and KoLee et al., p. 97, pl. 1, figs. 6–7.
2000Eoconodontus notchpeakensis (Miller); Reference Pyle and BarnesDubinina, p. 192, pl. 7, figs. 8–12, 16–20, ?figs. 14–15, pl. 8, figs. 2–7, 14, ?fig. 1, pl. 9, figs. 11–13.
2001Eoconodontus notchpeakensis (Miller); Reference LeeLee, p. 451, fig. 7 (17, 19, 24).
2002Eoconodontus notchpeakensis (Miller); Lee, p. 165, pl. 2, figs. 28, 29.
2002Eoconodontus notchpeakensis (Miller); Lee, p. 27, pl. 2, figs. 17, 19.
2003Eoconodontus notchpeakensis (Miller); Reference Heinsalu, Kaljo, Kurvits and ViiraHeinsalu et al., p. 147, text-figs. 9, 10.
2004cEoconodontus notchpeakensis (Miller); Reference Dong, Donoghue, Cheng and LiuDong et al., p. 1204, pl. 2, figs. 1, 3.
2004Eoconodontus notchpeakensis (Miller); Reference LeeLee, p. 60, fig. 5 (V–W).
2006Eoconodontus notchpeakensis (Miller); Reference Miller, Ethington, Evans, Holmer, Loch, Popov, Repetski, Ripperdan and TaylorMiller et al., p. 394, fig. 9 (O, P).
2007Eoconodontus notchpeakensis (Miller); Reference Landing, Westrop and KeppieLanding et al., p. 918, fig. 6 (q, r).
2011Eoconodontus notchpeakensis (Miller); Reference Miller, Evens, Freeman, Ripperdan and TaylorMiller et al., p. 604, fig. 7, K–N.
Description.—
Bimembrate apparatus of rounded and compressed elements. Both elements proclined to erect; basal cavity variable, from moderately large to very deep; cusp short to long, composed of white matter; surface smooth. Rounded element symmetrical or subsymmetrical, oval in cross section, possessing or lacking anterior and posterior keels. Compressed element asymmetrical due to lateral bending of the cusp and to the presence of carina on inner side of the base; cusp strongly compressed laterally, with prominent anterior and posterior keels.
Materials.—
226 specimens.
Occurrence.—
Known from the Wa’ergang section, where it ranges from the Eoconodontus Zone through the Cordylodus lindstromi Zone (upper part).
Genus Eodentatus Nicoll, Reference Nicoll1991
Type species.—
Eodentatus bicuspatus Nicoll and Shergold, Reference Nicoll and Shergold1991.
Eodentatus bicuspatus Nicoll and Shergold, Reference Nicoll and Shergold1991
1991Eodentatus bicuspatus Reference Nicoll and ShergoldNicoll and Shergold, p. 99, fig. 3.
Materials.—
Two specimens.
Occurrence.—
Known from the Wa’ergang section, where it occurs in the Cordylodus proavus Zone.
Remarks.—
The present specimens agree in all respects with those described and illustrated by Nicoll and Shergold (Reference Nicoll and Shergold1991). This species is characterized by ramiform elements with two denticles and an elongated ovate base.
Genus Granatodontus Chen and Gong, Reference Chen and Gong1986
Type species.—
Granatodontus ani (Wang, Reference Wang1985).
Granatodontus ani (Wang, Reference Wang1985)

Figure 18 (1–29) Granatodontus ani (Wang, Reference Wang1985); (1) Bitiao Formation, Wangcun section, GMPKU2652, lateral view; (2) Bitiao Formation, Wangcun section, GMPKU2653, lateral view; (3) Shenjiawan Formation, Wangcun section, GMPKU2654, lateral view; (4) Shenjiawan Formation, Wangcun section, GMPKU2655, lateral view; (5) Shenjiawan Formation, Wangcun section, GMPKU2656, lateral view; (6) Shenjiawan Formation, Wangcun section, GMPKU2657, posterior view; (7) Shenjiawan Formation, Wangcun section, GMPKU2658, lateral view; (8) Shenjiawan Formation, Wangcun section, GMPKU2659, lateral view; (9) Shenjiawan Formation, Wangcun section, GMPKU2660, lateral view; (10) Shenjiawan Formation, Wangcun section, GMPKU2661, lateral view; (11) Shenjiawan Formation, Wangcun section, GMPKU2662, lateral view; (12) Shenjiawan Formation, Wangcun section, GMPKU2663, lateral view; (13) Shenjiawan Formation, Wangcun section, GMPKU2664, lateral view; (14) Shenjiawan Formation, Wangcun section, GMPKU2665, lateral view; (15) Shenjiawan Formation, Wangcun section, GMPKU2666, lateral view; (16) Shenjiawan Formation, Wangcun section, GMPKU2667, lateral view; (17) Shenjiawan Formation, Wangcun section, GMPKU2668, lateral view; (18) Shenjiawan Formation, Wangcun section, GMPKU2669, lateral view; (19) Shenjiawan Formation, Wangcun section, GMPKU2670, lateral view; (20) Shenjiawan Formation, Wangcun section, GMPKU2671, lateral view; (21) Shenjiawan Formation, Wangcun section, GMPKU2672, lateral view; (22) Shenjiawan Formation, Wangcun section, GMPKU2673, lateral view; (23) Shenjiawan Formation, Wangcun section, GMPKU2674, lateral view; (24) Shenjiawan Formation, Wangcun section, GMPKU2675, lateral view; (25) Shenjiawan Formation, Wangcun section, GMPKU2676, lateral view; (26) Shenjiawan Formation, Wangcun section, GMPKU2677, lateral view; (27) Shenjiawan Formation, Wangcun section, GMPKU2678, lateral view; (28) Shenjiawan Formation, Wangcun section, GMPKU2679, lateral view; (29) Shenjiawan Formation, Wangcun section, GMPKU2680, lateral view. Relative scale bar represents 154 μm (1, 2, 5–24, 26, 28, 29), 144 μm (3, 4, 25), 164 μm (27).
1983Hirsutodontus sp. aff. Hirsutodontus primitivus Reference An, Zhang, Xiang, Zhang, Xu, Zhang, Jiang, Yang, Lin, Cui and YangAn et al., p. 104, pl. 4, figs. 22–23.
1985Hirsutodontus primitivus An et al., pl. 1, figs. 1–2.
1985Hirsutodontus? ani Wang in Chen et al., Reference Chen, Qian, Lin, Zhang, Wang, Yin and Erdtmann1985, p. 92, pl. 26, figs. 4–6.
1986Granatodontus ani (Wang); Reference Chen and GongChen and Gong, p. 149, pl. 26, fig. 8, pl. 27, figs. 1–5, 8–10, pl. 28, figs. 13, 14, pl. 31, fig. 10, text-fig. 53.
2000Hirsutodontus ani Wang; Reference Pyle and BarnesDubinina, p. 186, pl. 6, fig. 1a.
2002Granatodontus ani (Wang); Lee, p. 165, pl. 2, fig. 13.
2004Granatodontus ani (Wang); Reference LeeLee, p. 60, fig. 5 (P).
2004Granatodontus ani (Wang); Reference Wu and DongWu and Dong, p. 230, pl. 1, figs. 1–4.
2008Granatodontus ani (Wang); Lee, p. 228, fig. 1 (3–4), p. 229, pl. 1, fig. 1.
2008Granatodontus asymmetrica Lee, p. 229, pl. 1, figs. 2, 3.
2008Granatodontus muliticorrugata Lee, p. 229, pl. 1, figs. 4–6.
2009Granatodontus ani (Wang); Reference Lee, Lee, Miller and JeongLee et al., p. 423, fig. 7 (19), p. 425, fig. 8 (2).
Description.—
Coniform elements, erect to proclined and symmetrical, oval in cross section. Base large. Basal cavity large and deep, with apex extending beyond the point of flexure of cusp. The surface of the whole element is ornamented with numerous tiny nodes, rarely with a few tiny spines, except for the apex.
Materials.—
80 specimens.
Occurrence.—
Known from Wangcun section where it ranges from the Westergaardodina cf. calix-Prooneotodus rotundatus Zone through the Eoconodontus Zone.
Remarks.—
Chen and Gong (Reference Chen and Gong1986) suggested that Granatodontus ani (Wang, Reference Wang1985) is a paraconodont, based on its large basal cavity and lack of white matter. However, histological investigation verifies G. ani is a euconodont (Wu and Dong, Reference Wu and Dong2004; Murdock et al., Reference Murdock, Dong, Repetski, Marone, Stampanon and Donoghue2013). Conodont elements with conspicuous surface ornamentation, such as spines or nodes, should all be considered to be euconodonts rather than paraconodonts. This generalization can be made without histological study, based on a hypothesis proposed by Bengtson (Reference Bengtson1976).
Genus Hirsutodontus Miller, Reference Miller1969
Type species.—
Hirsutodontus hirsutus Miller, Reference Miller1969.
Hirsutodontus nodus (Zhang and Xiang in An et al., Reference An, Zhang, Xiang, Zhang, Xu, Zhang, Jiang, Yang, Lin, Cui and Yang1983)

Figure 19 (1–3) Gumella cuneata Müller and Hinz, Reference Müller and Hinz1991; (1) Huaqiao Formation, Wangcun section, GMPKU2681, lateral view; (2) Huaqiao Formation, Wangcun section, GMPKU2682, lateral view; (3) Chefu Formation, Wangcun section, GMPKU2683, lateral view; (4–13) Hirsutodontus nodus (Zhang and Xiang, 1983); (4) Shenjiawan Formation, Wangcun section, GMPKU2684, lateral view; (5) Shenjiawan Formation, Wangcun section, GMPKU2685, lateral view; (6) Shenjiawan Formation, Wangcun section, GMPKU2686, lateral view; (7) Shenjiawan Formation, Wangcun section, GMPKU2687, lateral view; (8) Shenjiawan Formation, Wangcun section, GMPKU2688, lateral view; (9) Shenjiawan Formation, Wa’ergang section, GMPKU2689, lateral view; (10) Shenjiawan Formation, Wa’ergang section, GMPKU2690, lateral view; (11) Shenjiawan Formation, Wa’ergang section, GMPKU2691, lateral view; (12) Panjiazui Formation, Wa’ergang section, GMPKU2692, lateral view; (13) Panjiazui Formation, Wa’ergang section, GMPKU2693, lateral view; (14–16, 22–24) Huayuanodontus tricornis (Dong, Reference Dong1993); (14) Chefu Formation, Wangcun section, GMPKU2242, posterior view; (15) Chefu Formation, Wangcun section, GMPKU2694, posterior view; (16) Chefu Formation, Wangcun section, GMPKU2695, posterior view; (22) Chefu Formation, Paibi section, GMPKU2696, posterior view; (23) Chefu Formation, Paibi section, GMPKU2245, posterior view; (24) close-up of (23) posterior view; (17–19) Hispidodontus resimus Nicoll and Shergold, Reference Nicoll and Shergold1991; (17) Bitiao Formation, Wangcun section, GMPKU2697, posterolateral view; (18) Bitiao Formation, Wangcun section, GMPKU2698, broken specimen, outer lateral view; (19) Bitiao Formation, Wangcun section, GMPKU2699, anterolateral view; (20, 21, 25, 26) Lugnathus hunanensis n. gen. n. sp.; (20) Panjiazui Formation, Wa’ergang section, GMPKU2700, lateral view; (21) Panjiazui Formation, Wa’ergang section, GMPKU2701, lateral view; (25) Panjiazui Formation, Wa’ergang section, holotype, GMPKU2702, lateral view; (26) Panjiazui Formation, Wa’ergang section, GMPKU2703, lateral view. Relative scale bar represents 502 μm (1), 269 μm (2), 270 μm (3), 164 μm (4–8, 10–13, 17, 18, 20, 21, 25, 26), 157 μm (9, 14, 19, 23), 146 μm (15, 16), 147 μm (22), 111 μm (24).
1983Teridontus nakamurai nodus Zhang and Xiang in An et al., p. 157, pl. 6, figs. 7, 8, text-fig. 14 (19).
1986Dasytodus nodus (Zhang and Xiang); Reference Chen and GongChen and Gong, p. 135, pl. 28, fig. 10, pl. 31, fig. 9, text-fig. 42.
1991Hirsutodontus nodus (Zhang and Xiang); Reference Nicoll and ShergoldNicoll and Shergold, figs. 4, 5 (1).
2001Hirsutodontus nodus (Zhang and Xiang); Reference LeeLee, p. 449, fig. 6 (19, 26), p. 451, fig. 7 (3).
2002Teridontus nodus (Zhang and Xiang); Lee, p. 27, pl. 2, fig. 20.
2002Hirsutodontus nodus (Zhang and Xiang); Lee, p. 165, pl. 2, fig. 14.
2004Hirsutodontus nodus (Zhang and Xiang); Reference LeeLee, p. 60, fig. 5 (S, X).
Materials.—
20 specimens.
Occurrence.—
Known from the Wangcun and Wa’ergang sections, where it occurs in the Eoconodontus Zone.
Remarks.—
An et al. (Reference An, Zhang, Xiang, Zhang, Xu, Zhang, Jiang, Yang, Lin, Cui and Yang1983) erected a new subspecies Teridontus nakamurai nodus Zhang and Xiang. Chen and Gong (Reference Chen and Gong1986) assigned this subspecies to Dasytodus. Having examined the published material and the new material from Hunan, we agree with Nicoll and Shergold (Reference Nicoll and Shergold1991) that T. nakamurai nodus should be assigned to Hirsutodontus rather than Dasytodus. The morphological difference between Dasytodus transmutatus and Hirsutodontus nodus has been fully elucidated by Nicoll and Shergold (Reference Nicoll and Shergold1991). The surface of the element of H. nodus is ornamented with only a few tiny nodes, so it differs from Hirsutodontus hirsutus Miller.
Genus Hispidodontus Nicoll and Shergold, Reference Nicoll and Shergold1991
Type species.—
Hispidodontus discretus Nicoll and Shergold, Reference Nicoll and Shergold1991.
Hispidodontus resimus Nicoll and Shergold, Reference Nicoll and Shergold1991
1991Hispidodontus resimus Reference Nicoll and ShergoldNicoll and Shergold, p. 107, figs. 14, 15, 16 (1–2).
Materials.—
Three specimens.
Occurrence.—
Known from the Wangcun section, where it occurs in the Westergaardodina cf. calix-Prooneotodus rotundatus Zone.
Remarks.—
The specimens at hand agree well with Hispidodontus resimus described and figured by Nicoll and Shergold (Reference Nicoll and Shergold1991). They are all S elements.
Genus Iapetognathus Landing, 1982
Type species.—
Pravognathus aengensis Lindström, Reference Lindström1955.
Iapetognathus aengensis (Lindström Reference Lindström1955)

Figure 20 (1–8) Lugnathus hunanensis n. gen. n. sp.; (1) Panjiazui Formation, Wa’ergang section, GMPKU2704, lateral view; (2) Panjiazui Formation, Wa’ergang section, GMPKU2705, lateral view; (3) Panjiazui Formation, Wa’ergang section, GMPKU2706, lateral view; (4) Panjiazui Formation, Wa’ergang section, GMPKU2707, lateral view; (5) Panjiazui Formation, Wa’ergang section, GMPKU2708, lateral view; (6) Panjiazui Formation, Wa’ergang section, GMPKU2709, lateral view; (7) Panjiazui Formation, Wa’ergang section, GMPKU2710, lateral view; (8) Panjiazui Formation, Wa’ergang section, GMPKU2711, lateral view; (9, 10) Hunanognathus tricuspidatus Dong, Reference Dong1993; (9) Huaqiao Formation, Wangcun section, GMPKU2712, posterior view; (10) Huaqiao Formation, Wangcun section, GMPKU2141, posterior view; (11–17) Iapetognathus aengesis (Lindström, Reference Lindström1955); (11) Panjiazui Formation, Wa’ergang section, GMPKU2713, S element, aboral view; (12) The same specimen, anterior view; (13) Panjiazui Formation, Wa’ergang section, GMPKU2714, S element, anterior view; (14) Panjiazui Formation, Wa’ergang section, GMPKU2715, S element, aboral view; (15) The same specimen, anterior view; (16) Panjiazui Formation, Wa’ergang section, GMPKU2716, S element, oblique aboral-posterior view; (17) Panjiazui Formation, Wa’ergang section, GMPKU2717, S element, anterior view; (18–24) Iapetognathus fluctivagus Nicoll, Miller, Nowlan, Repetski and Ethington, Reference Nicoll, Miller, Nowlan, Repetski and Ethington1999; (18) Panjiazui Formation, Wa’ergang section, GMPKU2718, P element, anterior view; (19) the same specimen, oral view; (20) Panjiazui Formation, Wa’ergang section, GMPKU2719, P element, oblique aboral-posterior view; (21) the same specimen, oblique lateral view; (22) the same specimen, oblique lateral view; (23) Panjiazui Formation, Wa’ergang section, GMPKU2720, P element, outer lateral view; (24) the same specimen, oblique anterior view; (25–28) Iapetognathus jilinensis Nicoll, Miller, Nowlan, Repetski and Ethington, Reference Nicoll, Miller, Nowlan, Repetski and Ethington1999; (25) Panjiazui Formation, Wa’ergang section, GMPKU2721, S element, oblique aboral-posterior view; (26) the same specimen, posterior view; (27) the same specimen, oblique oral-lateral view; (28) close-up of (27) showing cusp cross-section. Relative scale bar represents 139 μm (1–9, 11, 27), 123 μm (10), 72 μm (28).
1955Pravognathus aengensis Reference LindströmLindström, p. 585, pl. 5. figs. 10–13.
1982Iapetognathus preaengensis Landing in Reference Fortey, Landing and SkevingtonFortey et al., p. 124–126, text-figs. 6 (B, C), 8 (B, C, H).
1986Iapetognathus preaengensis Landing; Reference Landing, Barnes and StevensLanding, Barnes, and Stevens, p. 1934, pl. 1, fig. 8.
1986Iapetognathus sp. Reference Kalio, Borovko, Heinsalu, Hazanovits, Mens, Popov, Sergejeva, Sobolevskaja and ViiraKaljo et al., pl. 3, fig. 13.
1987Iapetognathus aengensis (Lindström); Reference OlgunOlgun, p. 60, fig. 30.
1988Iapetognathus preaengensis Landing; Reference BarnesBarnes, fig. 13cc only (not fig. 13 y, z, aa, bb, dd, ee). (part)
1988Iapetognathus preaengensis Landing; Reference Kalio, Heinsalu, Mens, Puura and ViiraKaljo et al., figs. 4f, 5f.
1988Iapetognathus sp. Reference Kalio, Heinsalu, Mens, Puura and ViiraKaljo et al., figs. 4bb, 5bb only.
1993Iapetognathus preaengensis Landing; Reference LandingLanding, p. 11, fig. 8 (1, ?2).
1993Iapetognathus sp. aff. I. preaengensis Landing; Reference Fåhræus and RoyFåhraeus and Roy, pl. 2, fig. 26 only.
1996Iapetognathus aengensis (Lindström); Reference LöfgrenLöfgren, p. 173, fig. 8 (G, K).
1999Iapetognathus aengensis (Lindström); Reference Nicoll, Miller, Nowlan, Repetski and EthingtonNicoll et al., p. 44, pl. 1, figs. 1–5, pl. 2, figs. 1–4, pl. 3, figs. 1–4, pl. 4, figs. 1–3, pl. 5, figs. 1–3.
2003Iapetognathus aengensis (Lindström); Reference Heinsalu, Kaljo, Kurvits and ViiraHeinsalu et al., p. 147, text-fig. 5 (26–30).
2012Iapetognathus aengensis (Lindström); Reference Terfelt, Bagnoli and StougeTerfelt et al., p. 230, fig. 2, A–E.
Materials.—
Five specimens.
Occurrence.—
Known only from the Wa’ergang section, where it occurs in the Cordylodus angulatus Zone (lower part).
Remarks.—
Nicoll et al. (Reference Nicoll, Miller, Nowlan, Repetski and Ethington1999) diagnosed multimembrate apparatus in which six element types have been identified, i.e., Sc, Sb, Sd, Xb, Pb, and Pa elements. However, only five elements of Iapetognathus aengensis have been recovered from Hunan, and all are S elements.
Iapetognathus fluctivagus Nicoll et al., Reference Nicoll, Miller, Nowlan, Repetski and Ethington1999
1970New Genus A Reference MillerMiller, p. 110, pl. 1, figs. 32–34, text-fig. 11E.
1982New Genus 2 (cordylodid) Reference Miller, Taylor, Stitt, Ethington, Hintze and TaylorMiller et al., text-figs. 5, 6.
1987Iapetognathus preaengesis Landing; Reference MillerMiller, fig. 3.
1988Iapetognathus preaengesis Landing; Reference BarnesBarnes, fig. 13z, aa only.
1995Iapetognathus n. sp. Reference Taylor and RepetskiTaylor and Repetski, p. 135.
1995Iapetognathus n. sp. 1 Reference Miller and StittMiller and Stitt, p. 108.
1995Iapetognathus n. sp. 1 Reference Miller and TaylorMiller and Taylor, p. 111.
1999Iapetognathus fluctivagus Reference Nicoll, Miller, Nowlan, Repetski and EthingtonNicoll et al., p. 46, pl. 6, figs. 1–5, pl. 7, figs. 1–4, pl. 8, figs. 1–2, pl. 9, figs. 1–7, pl. 10, figs. 1–6, pl. 11, figs. 1–6.
2004cIapetognathus fluctivagus Nicoll et al.; Reference Dong, Donoghue, Cheng and LiuDong et al., p. 1205, pl. 3, figs. 1–3.
non2007 Iapetognathus fluctivagus Nicoll et al.; Reference Pyle, Barnes and McanallyPyle et al., 1656, p. 1733, fig. 10 (8, 9).
2012Iapetognathus? fluctivagus Nicoll et al.; Reference Terfelt, Bagnoli and StougeTerfelt et al., p. 230, fig. 2, F–G, H–K.
Materials.—
Three specimens.
Occurrence.—
Known only from the Wa’ergang section, where it occurs in the upper part of the Cordylodus lindstromi Zone.
Remarks.—
Although Iapetognathus fluctivagus is widespread in North America, specimens of it are very rare in China. So far, only three specimens belonging to P elements have been recovered in western Hunan, South China, by means of sampling and processing on an extremely large scale. Cooper et al. (Reference Cooper, Nowlan and Williams2001) established a stratotype for the base of the Ordovician based on the FAD of this species at Green Point in Newfoundland, Canada.
Iapetognathus jilinensis Nicoll et al., Reference Nicoll, Miller, Nowlan, Repetski and Ethington1999
1995Iapetognathus n. sp. A Reference Nowlan and NicollNowlan and Nicoll, p. 115–116.
1999Iapetognathus jilinensis Reference Nicoll, Miller, Nowlan, Repetski and EthingtonNicoll et al., p. 48, pl. 12, figs. 1a–4f, pl. 13, figs. 1a–3f, pl. 14, figs. 1a–4b.
Materials.—
Only one specimen.
Occurrence.—
Known only from the Wa’ergang section, where it occurs in the Cordylodus lindstromi Zone (Upper Part).
Remarks.—
Only a single specimen belonging to S element has been recovered from western Hunan.
Genus Mamillodus Abaimova, Reference Abaimova1983
Type species.—
Mamillodus tubularis Abaimova, Reference Abaimova1983.
Mamillodus ruminatus Dubinina, Reference Dubinina2000

Figure 21 (1, 2) Laiwugnathus hunanensis n. sp.; (1) Huaqiao Formation, Wangcun section, holotype, GMPKU2722, posterior view; (2) Huaqiao Formation, Wangcun section, GMPKU2723, posterior view; (3) Laiwugnathus cf. kouzhenensis An, Reference An1982, Chefu Formation, Wangcun section, GMPKU2724, posterior view; (4–6) Laiwugnathus transitans n. sp.; (4) Chefu Formation, Wangcun section, GMPKU2725, posterior view; (5) Chefu Formation, Wangcun section, holotype, GMPKU2726, posterior view; (6) Bitiao Formation, Wangcun section, GMPKU2727, lateral view; (7) Mamillodus ruminatus Dubinina, Reference Dubinina2000, Shenjiawan Formation, Wa’ergang section, GMPKU2728, posterior view; (8–12, 15, 16) Miaognathus multicostatus n. gen. n. sp.; (8) Shenjiawan Formation, Wa’ergang section, GMPKU2729, lateral view; (9) Shenjiawan Formation, Wa’ergang section, GMPKU2730, lateral view; (10) Shenjiawan Formation, Wa’ergang section, GMPKU2731, lateral view; (11) Shenjiawan Formation, Wa’ergang section, GMPKU2732, lateral view; (12) Shenjiawan Formation, Wa’ergang section, holotype, GMPKU2733, lateral view; (15) Shenjiawan Formation, Wa’ergang section, GMPKU2734, lateral view; (16) Shenjiawan Formation, Wa’ergang section, GMPKU2735, lateral view; (13, 14, 17–19) Millerodontus intermedius n. gen. n. sp.; (13) Shenjiawan Formation, Wa’ergang section, GMPKU2736, lateral view; (14) Shenjiawan Formation, Wa’ergang section, GMPKU2737, lateral view; (17) Shenjiawan Formation, Wa’ergang section, holotype, GMPKU2252, lateral view; (18) the same specimen, posterior-lateral view; (19) Shenjiawan Formation, Wa’ergang section, GMPKU2167, lateral view; (20–23) Monocostodus sevierensis Miller, Reference Miller1980; (20) Panjiazui Formation, Wa’ergang section, GMPKU2738, lateral view; (21) Panjiazui Formation, Wa’ergang section, GMPKU2739, lateral view; (22) Panjiazui Formation, Wa’ergang section, GMPKU2740, lateral view; (23) Panjiazui Formation, Wa’ergang section, GMPKU2741, lateral view; Relative scale bar represents 146 μm (1, 17, 23), 156 μm (2–16, 18, 20–22), 103 μm (19).
2000 Mamillodus ruminatus Reference DubininaDubinina, p. 171, pl. 7, figs. 1–6.
Materials.—
Only one specimen.
Occurrence.—
Known from the Wa’ergang section, where it occurs in the Eoconodontus Zone.
Remarks.—
The specimens at hand agree closely with Mamillodus ruminatus. Dubinina indicated it is a paraconodont. Based on its surface ornamentation, however, it is probably a euconodont (Bengtson, Reference Bengtson1976).
Genus Millerodontus new genus
Type species.—
Millerodontus intermedius n. gen. n. sp.
Diagnosis.—
Coniform euconodont elements, proclined, symmetrical, with rounded cross section. Keels or costae are lacking. Basal cavity is very deep.
Etymology.—
In honor of James F. Miller, Missouri State University, USA, the name is composed from Miller and a derivative of Greek, odon, tooth.
Millerodontus intermedius new genus new species
Figures 21.13, 21.14, 21.17–21.19, 22.1–22.8

Figure 22 Conodont images using oil immersion techniques (Dong et al., 2005c) with Differential Interference Contrast (Nomarski) illumination, showing euconodont structure. (1–8) Millerodontus intermedius n. gen. n. sp.; (1–4) The same specimen as Figure 21.17; (1) overview, showing the crown structure of euconodont; (2) close-up of the cusp of (1); (3) close-up of the tip of (2), showing the regeneration phenomena of euconodont; (4) close-up of of (3), showing the tiny phosphate crystals perpendicular to growth axis (shown by arrow); (5) the same specimen as Figure 21.14 overview, showing the crown structure of euconodont; (6–8) the same specimen as Figure 21.13; (6) overview; (7) close-up of of (6); (8) close-up of the tip of (7), showing the crown structure of euconodont. Relative scale bar represents 55 μm (1), 19 μm (2), 14 μm (3), 12 μm (4), 71 μm (5), 75 μm (6), 62 μm (7), 24 μm (8).
Diagnosis.—
As for genus.
Description.—
Moderate to small coniform elements, proclined, and symmetrical. The apex is pointed. Both anterior and posterior sides are rounded, without keels or costae. Basal cavity is very deep, the apex of which extends nearly to the tip of the element. The cross section is circular. Crown tissues and the phenomenon of regeneration could be observed at the apical portion of element.
Etymology.—
intermedius, Latin, intermediate.
Types.—
Holotype: GMPKU2252 from the Proconodontus tenuiserratus Zone, Shenjiawan Formation, Wa’ergang section, Wa’ergang village, Taoyuan county, Hunan Province.
Materials.—
10 specimens.
Occurrence.—
Known from the Wa’ergang and Wangcun sections, where it ranges from the Proconodontus tenuiserratus Zone through the Cordylodus proavus Zone.
Comparision.—
In gross morphology, Millerodontus intermedius n. gen. n. sp. resembles Prooneotodus rotundatus, but it is recognized as euconodont in the light of histological investigation. M. intermedius is a representative of the model of early evolution of euconodonts proposed by Bengtson (Reference Bengtson1976), according to which euconodont elements originated from paraconodonts through acquisition of a crown. M. intermedius is probably the transitional form between P. rotundatus and Proconodontus tenuiserratus.
Remarks.—
The majority of the specimens which resemble Prooneotodus rotundatus have thick sheaths, just like Miller’s interpretation that a thin layer of apatite covers a massive white basal cone (Miller, Reference Miller1980, p. 31–32). Not all specimens could be investigated by means of the oil immersion technique, and thin sectioning would ruin the specimens. Therefore, the number of specimens of Millerodontus intermedius n. gen. n. sp. is probably much more than ten.
Genus Monocostodus Miller, Reference Miller1980
Type species.—
Acodus sevierensis Miller, Reference Miller1969.
Monocostodus sevierensis (Miller, Reference Miller1969)
1969Acodus sevierensis Reference MillerMiller, p. 418, pl. 63, figs. 25–31 (not figs. 21–24, 32). (part)
1969Acontiodus (Semiacontiodus) unicostatus Reference MillerMiller, p. 421, pl. 64, figs. 19–54 (not figs. 46–48). (part)
1971Drepanodus simplex Branson and Mehl; Reference Druce and JonesDruce and Jones, p. 74, text-fig. 24b, pl. 13, figs. 1–4.
1973Semiacontiodus sevierensis (Miller), Lindström, p. 442–445.
1980Monocostodus sevierensis (Miller); Reference MillerMiller, p. 27, pl. 2, figs. 8–9, text-fig. 4U.
1982Monocostodus sevierensis (Miller); Landing in Reference Fortey, Landing and SkevingtonFortey et al., p. 123, text-fig. 8 (Q, R).
1983Monocostodus sevierensis (Miller); Reference An, Zhang, Xiang, Zhang, Xu, Zhang, Jiang, Yang, Lin, Cui and YangAn et al., p. 108, pl. 6, figs. 19–23.
1985Monocostodus sevierensis (Miller); Reference DongDong, p. 400, pl. 3, figs. 8, 11.
? 1985Monocostodus sevierensis (Miller); Reference WangWang in Chen et al., p. 93, pl. 25, figs. 9–11, text-fig. 14 (7).
1987Monocostodus sevierensis (Miller); Reference DongDong, p. 163, pl. 1, figs. 26, 32, pl. 2, figs. 1, 2.
1993Monocostodus sevierensis (Miller); Reference LandingLanding, p. 6, fig. 4 (10, 18, 19).
1996Drepanodus simplex Branson and Mehl; Reference Lee, Lee and KoLee et al., p. 97, pl. 1, fig. 17.
1998Monocostodus sevierensis (Miller); Reference Rao and TortelloRao and Tortello, p. 43, pl. 1, figs. 13–18.
2000Drepanodus simplex Branson and Mehl; Reference Zhao, Zhang and XiaoZhao et al., p. 198, pl. 9, figs. 1–9.
2000Monocostodus sevierensis (Miller); Reference Zhao, Zhang and XiaoZhao et al., p. 206, pl. 8, figs. 1–7.
2003Monocostodus sevierensis (Miller); Reference Landing, Westrop and van Aller HernickLanding et al., p. 86, fig. 7 (14, 18).
Description.—
Slender coniform elements, erect to reclined, with short base and long cusp. Cross section is circular through base and the aboral half of the cusp, but it becomes extremely laterally compressed through the oral half of cusp. The anterior and posterior sides of the oral half of cusp are keeled. Basal cavity is shallow. The slender cusp is composed entirely of white matter.
Materials.—
12 specimens.
Occurrence.—
Known from the Wa’ergang section, where it ranges from the Cordylodus lindstromi Zone (upper part) through the Cordylodus angulatus Zone.
Genus Proconodontus Miller, Reference Miller1969
Type species.—
Proconodontus muelleri Miller, Reference Miller1969.
Proconodontus muelleri Miller, Reference Miller1969

Figure 23 (1–6) Proacodus pulcherus (An, Reference An1982); (1) Bitiao Formation, Wangcun section, GMPKU2793, anterior view; (2) Bitiao Formation, Wangcun section, GMPKU2794, posterior view; (3) Bitiao Formation, Wangcun section, GMPKU2795, posterior view; (4) Shenjiawan Formation, Wa’ergang section, GMPKU2796, posterior view; (5) Bitiao Formation, Wangcun section, GMPKU2797, posterior view; (6) Shenjiawan Formation, Wa’ergang section, GMPKU2798, posterior view; (7) Proacodus cf. pulcherus (An, Reference An1982) Bitiao Formation, Wangcun section, GMPKU2799, posterior view; (8–32) Proconodontus muelleri Miller, Reference Miller1969; (8) Shenjiawan Formation, Wangcun section, GMPKU2800, lateral view; (9) Shenjiawan Formation, Wa’ergang section, GMPKU2801, lateral view; (10) Shenjiawan Formation, Wa’ergang section, GMPKU2802, lateral view; (11) Shenjiawan Formation, Wangcun section, GMPKU2803, lateral view; (12) Shenjiawan Formation, Wa’ergang section, GMPKU2804, lateral view; (13) Shenjiawan Formation, Wa’ergang section, GMPKU2805, lateral view; (14) Shenjiawan Formation, Wa’ergang section, GMPKU2806, lateral view; (15) Shenjiawan Formation, Wangcun section, GMPKU2807, lateral view; (16) Shenjiawan Formation, Wa’ergang section, GMPKU2808, lateral view; (17) Shenjiawan Formation, Wa’ergang section, GMPKU2809, lateral view; (18) Shenjiawan Formation, Wa’ergang section, GMPKU2810, lateral view; (19) Shenjiawan Formation, Wa’ergang section, GMPKU2811, lateral view; (20) Shenjiawan Formation, Wa’ergang section, GMPKU2812, lateral view; (21) Shenjiawan Formation, Wa’ergang section, GMPKU2813, lateral view; (22) Shenjiawan Formation, Wa’ergang section, GMPKU2814, lateral view; (23) Shenjiawan Formation, Wa’ergang section, GMPKU2815, lateral view; (24) Shenjiawan Formation, Wa’ergang section, GMPKU2816, lateral view; (25) Shenjiawan Formation, Wa’ergang section, GMPKU2817, lateral view; (26) Shenjiawan Formation, Wa’ergang section, GMPKU2818, lateral view; (27) Shenjiawan Formation, Wa’ergang section, GMPKU2819, lateral view; (28) Shenjiawan Formation, Wa’ergang section, GMPKU2820, lateral view; (29) Shenjiawan Formation, Wa’ergang section, GMPKU2821, lateral view; (30) Shenjiawan Formation, Wa’ergang section, GMPKU2822, lateral view; (31) Shenjiawan Formation, Wa’ergang section; GMPKU2823, lateral view; (32) Shenjiawan Formation, Wa’ergang section, GMPKU2824, lateral view. Relative scale bar represents 165 μm (1–6, 8–21, 23, 24, 28), 156 μm (7, 27), 173 μm (22, 25, 26, 29–32).
1969Proconodontus mülleri mülleri Reference MillerMiller, p. 437, text-fig. 5H, pl. 66, figs. 30–40.
1970Proconodontus mülleri mülleri Miller; Reference MillerMiller, pl. 2, fig. 18.
1971Proconodontus mulleri mulleri Miller; Reference Ethington and ClarkEthington and Clark, pl. 1, fig. 25.
1971Coelocerodontus burkei Reference Druce and JonesDruce and Jones, p. 61, text-fig. 22a (not text-fig. 22e), pl. 11, figs. 9–11 (not figs. 5–8, 12). (part)
? 1973Proconodontus muelleri Miller; Reference MüllerMüller, p. 42, pl. 3, figs. 4–7, ?10 (not fig. 8). (part)
1976Proconodontus muelleri Miller; Reference Miller and PadenMiller and Paden, p. 595, pl. 1, figs. 4, 5.
1978Proconodontus muelleri muelleri Miller; Reference Tipnis, Chatterton and LudvigsenTipnis et al., pl. 1, figs. 13, 17.
1980Proconodontus muelleri Miller; Reference MillerMiller, p. 29, text-fig. 4C, pl. 1, fig. 7.
1981Proconodontus muelleri Miller; Miller in Reference RobisonRobison, p. W146, fig. 95 (2a, b).
1982Proconodontus muelleri muelleri Miller; Reference Fortey, Landing and SkevingtonFortey et al., p. 125, text-fig. 9 (K).
1982Proconodontus muelleri Miller; Reference AnAn, p. 141, pl. 12, figs. 8, 9, 11–13, pl. 16, figs. 10, 12.
1983Proconodontus muelleri Miller; Reference An, Zhang, Xiang, Zhang, Xu, Zhang, Jiang, Yang, Lin, Cui and YangAn et al., p. 126, pl. 5, fig. 21 only (not figs. 15, 16, 23, 24). (part)
1985Proconodontus muelleri Miller; Reference NowlanNowlan, p. 114, fig. 5.1.
1985Proconodontus muelleri Miller; Reference WangWang, p. 231, pl. 2, figs. 16–20 (not fig. 21), pl. 6, figs. 17–19, pl. 7, figs. 10, 27, pl. 8, f’igs. 17, 18, pl. 12, fig. 7, pl. 13, figs. 1, 6. (part)
? 1985Proconodontus muelleri Miller; Reference WangWang, p. 231, pl. 9, figs. 16–18, pl. 13, fig. 25.
1986Proconodontus muelleri Miller; Reference Chen and GongChen and Gong, p. 159, pl. 32, figs. 1, 3, 10, 12–14, 17, pl. 33, figs. 3–5, 11 (not pl. 19, fig. 6), text-fig. 60 (1–2; 4–6; not 3). (part)
1987Proconodontus muelleri Miller; Reference DongDong, p. 169, pl. 1, figs. 3–5, text-figs. 3B, C, 4D.
1987Proconodontus muelleri Miller; Reference AnAn, p. 109, pl. 2, figs. 4, 17.
1987Proconodontus aff. muelleri Miller; Reference AnAn, p. 110, pl. 2, figs. 19–21.
1988Proconodontus muelleri Miller; Andres, p. 125, pl. 10, figs. 1–8, pl. 11, figs. 1–8, pl. 12, figs. 1–4, text-figs. 20–25.
1988Proconodontus muelleri Miller; Reference Lee and LeeLee and Lee, pl. 2, figs. 27–29.
1991Proconodontus muelleri Miller; Reference Müller and HinzMüller and Hinz, p. 56, pl. 42, figs. 1–16, text-figs. 22B, C.
1993Proconodontus muelleri Miller; Reference LandingLanding, p. 6, fig. 4 (5).
1996Proconodontus muelleri Miller; Reference Lee, Lee and KoLee et al., p. 97, pl. 1, fig. 3.
? 2000Proconodontus muelleri Miller; Reference Pyle and BarnesDubinina, p. 194, pl. 7, fig. 7.
2001Proconodontus muelleri Miller; Reference LeeLee, p. 451, fig. 7.15.
2002Proconodontus muelleri Miller; Lee, p. 165, pl. 2, fig. 26.
2002Proconodontus muelleri Miller; Lee, p. 27, pl. 2, fig. 15.
2004Proconodontus muelleri Miller; Reference LeeLee, p. 60, fig. 5 (H).
2006Proconodontus muelleri Miller; Reference Miller, Ethington, Evans, Holmer, Loch, Popov, Repetski, Ripperdan and TaylorMiller et al., p. 394, fig. 9 (S).
2007Proconodontus muelleri Miller; Reference Landing, Westrop and KeppieLanding et al., p. 918, fig. 6 (n, o).
2009Proconodontus muelleri Miller; Reference Lee, Lee, Miller and JeongLee et al., p. 423, fig. 7 (8).
2011Proconodontus muelleri Miller; Reference Miller, Evens, Freeman, Ripperdan and TaylorMiller et al., p. 604, fig. 7, I, J.
Description.—
Coniform elements, moderate to large, proclined and symmetrical or nearly so. The cross section is circular at the apical portion and it changes into an oval to moderately compressed outline toward the base. Basal cavity is very large and deep, with the apex extending nearly to the tip of it. Both anterior and posterior edges are characterized by sharp keels of variable extent. Usually, the posterior keel is more conspicuous and extends from the tip to the base, while the anterior keel extends from the tip to variable distances down toward the base of the element. Regeneration of the tip is often observed at the apical portion of the element.
Materials.—
200 specimens.
Occurrence.—
Known from the Wangcun and Wa’ergang sections, where it ranges from the Proconodontus Zone through the Eoconodontus Zone.
Remarks.—
Miller (Reference Miller1980) believed white matter is lacking in Proconodontus. However, Müller and Hinz (Reference Müller and Hinz1991) found white matter in the keel and in the regenerated part of the tip. Later, they verified this observation by histological investigation (Müller and Hinz, Reference Müller and Hinz1998, p. 108–110, fig. 18.3). Indeed, our histological study on many specimens of Proconodontus muelleri recovered from Hunan, South China also verified this conclusion (Xi-ping Dong and Philip C. J. Donoghue, unpublished data).
Proconodontus posterocostatus Miller, Reference Miller1980

Figure 24 (1, 2) Proconodontus posterocostatus Miller, Reference Miller1980; (1) Shenjiawan Formation, Wa’ergang section, GMPKU2825, lateral view; (2) Shenjiawan Formation, Wa’ergang section, GMPKU2826, lateral view; (3–6) Proconodontus serratus Miller, Reference Miller1969; (3) Shenjiawan Formation, Wa’ergang section, GMPKU2827, lateral view; (4) Shenjiawan Formation, Wa’ergang section, GMPKU2828, lateral view; (5) Shenjiawan Formation, Wa’ergang section, GMPKU2829, lateral view; (6) Shenjiawan Formation, Wa’ergang section, GMPKU2830, lateral view; (7, 8, 11) Proconodontus tenuiserratus Miller, Reference Miller1980; (7) Shenjiawan Formation, Wa’ergang section, GMPKU2831, lateral view; (8) Shenjiawan Formation, Wa’ergang section, GMPKU2832, lateral view; (11) Shengjiawan Formation, Wangcun section, GMPKU2168, lateral view; (9, 10, 12–32) Prooneotodus gallatini Müller, Reference Müller1959; (9) Bitiao Formation, Wangcun section, GMPKU2833, lateral view; (10) Bitiao Formation, Wangcun section, GMPKU2834, lateral view; (12) Bitiao Formation, Wangcun section, GMPKU2835, lateral view; (13) Bitiao Formation, Wangcun section, GMPKU2836, lateral view; (14) Bitiao Formation, Wangcun section, GMPKU2837, lateral view; (15) Bitiao Formation, Wangcun section, GMPKU2838, lateral view; (16) Bitiao Formation, Wangcun section, GMPKU2839, lateral view; (17) Bitiao Formation, Wangcun section, GMPKU2840, lateral view; (18) Bitiao Formation, Wangcun section, GMPKU2841, lateral view; (19) Bitiao Formation, Wangcun section, GMPKU2842, lateral view; (20) Bitiao Formation, Wangcun section, GMPKU2843, lateral view; (21) Bitiao Formation, Wangcun section, GMPKU2844, lateral view; (22) Bitiao Formation, Wangcun section, GMPKU2845, lateral view; (23) Bitiao Formation, Wangcun section, GMPKU2846, lateral view; (24) Bitiao Formation, Wangcun section, GMPKU2847, lateral view; (25) Bitiao Formation, Wangcun section, GMPKU2848, lateral view; (26) Bitiao Formation, Wangcun section, GMPKU2849, lateral view; (27) Bitiao Formation, Wangcun section, GMPKU2850, lateral view; (28) Bitiao Formation, Wangcun section, GMPKU2851, lateral view; (29) Bitiao Formation, Wangcun section, GMPKU2852, lateral view; (30) Bitiao Formation, Wa’ergang section, GMPKU2853, lateral view; (31) Bitiao Formation, Wa’ergang section, GMPKU2854, lateral view; (32) Shenjiawan Formation, Wa’ergang section, GMPKU2855, lateral view. Scale bar represents 138 μm (1–32).
1971Coelocerodontus burkei Reference Druce and JonesDruce and Jones, p. 61, pl. 11, figs. 7, 8 (not figs. 5, 6, 9–12). (part)
1980Proconodontus posterocostatus Reference MillerMiller, p. 30, text-fig. 4B, pl. 1, figs. 4–6.
non1982 Proconodontus posterocostatus Miller; Reference AnAn, p. 142, pl. 12, figs. 3, 4, 6, 7, 10.
1986Proconodontus posterocostatus Miller; Reference Chen and GongChen and Gong, p. 161, pl. 25, figs. 1, 6, pl. 30, figs. 1, 3–4, 8–13, pl. 32, fig. 8, pl. 33, figs. 7, 9 (not pl. 19, fig. 16), text-fig. 61 (1, 3–8, not 2). (part)
2002Proconodontus posterocostatus Miller; Lee, p. 165, pl. 2, fig. 25.
2002Proconodontus posterocostatus Miller; Lee, p. 27, pl. 2, fig. 12.
2004cProconodontus posterocostatus Miller; Reference Dong, Donoghue, Cheng and LiuDong et al., pl. 1, fig. 3.
2004cProconodontus serratus Miller; Reference Dong, Donoghue, Cheng and LiuDong et al., pl. 1, fig. 11.
? 2009Proconodontus posterocostatus Miller; Reference Lee, Lee, Miller and JeongLee et al., p. 423, fig. 7 (7, 9).
Description.—
Moderate to large coniform elements, erect to proclined and symmetrical or nearly so. The cross section is circular at the apical portion and changes to an oval outline toward the base. Basal cavity is very large and deep, the apex extends nearly to the tip of the element; posterior edges are characterized by a sharp keel that extends from apex to a variable distance down the posterior edge of the element and may be one-third of the total length of the element; anterior keel is lacking.
Materials.—
Five specimens.
Occurrence.—
Known from the Wa’ergang sections, where it ranges from the Proconodontus Zone through the Eoconodontus Zone.
Proconodontus serratus Miller, Reference Miller1969
1969Proconodontus muelleri serratus Reference MillerMiller, p. 438, pl. 66, figs. 41–44.
1971Coelocerodontus burkei Reference Druce and JonesDruce and Jones, p. 61, pl. 11, fig. 12 (not figs. 5–11). (part)
1973Proconodontus muelleri serratus Miller; Lindström, p. 409, pl. 1, fig. 6.
1973Proconodontus serratus Miller; Reference MüllerMüller, p. 44, pl. 4, figs. 1, 2.
1980Proconodontus serratus Miller; Reference MillerMiller, p. 31, pl. 1, fig. 13, text-fig. 4D.
1980Proconodontus serratus Miller; Reference Landing, Ludvigen and von BitterLanding et al., p. 33, fig. 8I–L.
1982Proconodontus muelleri serratus Miller; Reference Fortey, Landing and SkevingtonFortey et al., p. 124, fig. 9 (L).
1983Proconodontus muelleri Miller; Reference An, Zhang, Xiang, Zhang, Xu, Zhang, Jiang, Yang, Lin, Cui and YangAn et al., p. 126, pl. 5, fig. 24a, b. (part)
1985Proconodontus serratus Miller; Reference WangWang, p. 233, pl. 6, fig. 26.
1986Proconodontus serratus Miller; Reference Chen and GongChen and Gong, p. 162, pl. 33, figs. 1, 6, 10, text-fig. 62.
1988Proconodontus serratus Miller; Reference Lee and LeeLee and Lee, pl. 2, figs. 30, 31.
1991Proconodontus serratus Miller; Reference Müller and HinzMüller and Hinz, p. 56, pl. 42, figs. 17–21, pl. 43, figs. 1–3, 6, 7, text-fig. 22A.
2000Proconodontus serratus Miller; Reference Pyle and BarnesDubinina, p. 195, pl. 6, figs. 21, 21a.
2006Proconodontus serratus Miller; Reference Miller, Ethington, Evans, Holmer, Loch, Popov, Repetski, Ripperdan and TaylorMiller et al., p. 394, fig. 9 (Q, R).
2007Proconodontus serratus Miller; Reference Landing, Westrop and KeppieLanding et al., p. 918, fig. 6 (t).
2011Proconodontus serratus Miller; Reference Miller, Evens, Freeman, Ripperdan and TaylorMiller et al., p. 604, fig. 7C, D.
Description.—
Moderate to large coniform elements, proclined and symmetrical. The cross section is laterally compressed. Basal cavity is very large and deep, with the apex extending nearly to the tip of the element. Anterior and posterior edges are characterized by prominent keels. The posterior keel is serrated except for the apical portion.
Materials.—
Six specimens.
Occurrence.—
Known from the Wa’ergang sections, where it ranges from the Proconodontus Zone through the Eoconodontus Zone.
Proconodontus tenuiserratus Miller, Reference Miller1980
1971Coelcerodontus burkei Reference Druce and JonesDruce and Jones, p. 61, text-fig. 22e, pl. 11, figs. 5, 6 (not figs. 7–12). (part)
1980Proconodontus tenuiserratus Reference MillerMiller, p. 31, text-fig. 4A, pl. 1, figs. 1–3.
1982Proconodontus tenuiserratus Miller; Reference AnAn, p. 143, pl. 12, figs. 1, 2, 5.
1985Proconodontus tenuiserratus Miller; Reference WangWang, p. 233, pl. 8, fig. 7, ?pl. 12, fig. 8.
1986Proconodontus tenuiserratus Miller; Reference Chen and GongChen and Gong, pl. 29, figs. 1–16, pl. 30, figs. 2, 5–7, text-fig. 63.
2000Proconodontus tenuiserratus Miller; Reference Pyle and BarnesDubinina, p. 195, pl. 6, fig. 22.
2002aProconodontus tenuiserratus Miller; Reference LeeLee, p. 165, pl. 2, fig. 24.
2002bProconodontus tenuiserratus Miller; Reference LeeLee, p. 27, pl. 2, figs. 10, 11.
2004Proconodontus tenuiserratus Miller; Reference LeeLee, p. 60, fig. 5 (Y).
2004cProconodontus tenuiserratus Miller; Reference Dong, Donoghue, Cheng and LiuDong et al., pl. 1, figs. 7, 12, pl. 2, fig. 26.
Description.—
Moderate to small coniform elements, erect to proclined and symmetrical. The cross section is usually circular, rarely compressed laterally near the base. Basal cavity is very deep, with the apex extending nearly to the tip of the element. The posterior side is characterized by a keel that is small, finely serrated, and extends from tip about halfway to the base, but it extends from the tip nearly to the base in some advanced forms. Anterior keel is lacking.
Materials.—
12 specimens.
Occurrence.—
Known from the Wa’ergang section, where it ranges from the Proconodontus tenuiserratus Zone through the Proconodontus Zone.
Genus Teridontus Miller, Reference Miller1980
Type species.—
Oneotodus nakamurai Nogami, Reference Nogami1967.
Teridontus nakamurai (Nogami, Reference Nogami1967)
Figure 25.4, 25.5, 25.8–25.15, 25.17–25.22
1959Oneotodus sp. A Reference MüllerMüller, p. 458, pl. 13, fig. 17.
1967Oneotodus nakamurai Reference NogamiNogami, p. 216, text-figs. 3A, B, C, ?D, ?E, pl. 1, figs. 9–13.
1969Oneotodus nakamurai Nogami; Reference MillerMiller, p. 435, text-fig. 5E, pl. 63, figs. 1–10.
1971Oneotodus nakamurai Nogami; Reference Druce and JonesDruce and Jones, p. 82, text-fig. 26i, pl. 10, figs. 3, 4, 7, 8. (part)
1973Oneotodus nakamurai Nogami; Reference MüllerMüller, p. 41, pl. 5, fig. 4.
1971Oneotodus datsonensis Reference Druce and JonesDruce and Jones, p. 80, text-fig. 26c, pl. 14, figs. 1–3. (part)
1980Teridontus nakamurai (Nogami); Reference MillerMiller, p. 34, pl. 2, figs. 15, 16, text-fig. 4 (O).
1982Teridontus nakamurai (Nogami); Reference AnAn, p. 150, pl. 14, figs. 1–11, pl. 15, fig. 11.
1983Teridontus nakamurai (Nogami); Reference An, Zhang, Xiang, Zhang, Xu, Zhang, Jiang, Yang, Lin, Cui and YangAn et al., p. 156, pl. 6, figs. 1–6.
1985Teridontus nakamurai (Nogami); Reference WangWang, p. 241, pl. 3, figs. 1, 2, 4, pl. 6, figs. 9–12, pl. 8, figs. 1–3, pl. 10, figs. 1–3, pl. 13, figs. 5, 11, 12.
1985Teridontus nakamurai (Nogami); Reference DongDong, p. 405, pl. 1, figs. 5, 18, text-fig. 1 (H).
1986Teridontus nakamurai (Nogami); Reference Chen and GongChen and Gong, p. 192, pl. 34, fig. 12, pl. 39, figs. 1, 3, 9, 11–13, pl. 40, figs. 1, 3, 5, 7, 9, 12–13, pl. 47, figs. 7, 8, pl. 48, fig. 16, text-fig. 79 (1–12, 15, 16).
1987Teridontus nakamurai (Nogami); Reference DongDong, p. 175, pl. 1, figs. 27–28.
1996Teridontus nakamurai (Nogami); Reference Lee, Lee and KoLee et al., p. 97, pl. 1, figs. 2, 8.
1999Teridontus nakamurai (Nogami); Reference Parsons and ClarkParsons and Clark, p. 7, fig. 5 (17–18, ?23).
2000Teridontus nakamurai (Nogami); Reference Pyle and BarnesDubinina, p. 196, pl. 6, figs. 2, 4, 5, 7, 8, 8a.
2000Teridontus nakamurai (Nogami); Reference Zhao, Zhang and XiaoZhao et al., p. 227, pl. 9, figs. 19–26.
2001Teridontus nakamurai (Nogami); Reference LeeLee, p. 450, fig. 7 (1, 2, 4–8, 10–13).
2002Teridontus nakamurai (Nogami); Lee, p. 165, pl. 2, fig. 15.
2002Teridontus nakamurai (Nogami); Lee, p. 27, pl. 2, fig. 21.
2003Teridontus nakamurai (Nogami); Reference Landing, Westrop and van Aller HernickLanding et al., p. 86, fig. 7 (3).
2005Teridontus nakamurai (Nogami); Reference Ortega and AlbanesiOrtega and Albanesi, p. 365, figs. 17–18.
2007Teridontus nakamurai (Nogami); Reference Landing, Westrop and KeppieLanding et al., p. 918, fig. 6 (i–l).
2007Teridontus nakamurai (Nogami); Reference Pyle, Barnes and McanallyPyle et al., p. 1733, fig. 10 (6, 7).
2009Teridontus nakamurai (Nogami); Reference Lee, Lee, Miller and JeongLee et al., p. 423, fig. 7 (2).
Description.—
Simple coniform elements with much morphological variation. Base is usually long but is short in advanced forms. Cusp is erect to reclined and is almost entirely composed of white matter that terminates in sharp boundary that is transverse to the cusp at a point slightly above the tip of basal cavity. Cross section is circular to oval. Fine lengthwise striae are usually preserved on the surface of the whole element.
Materials.—
100 specimens.
Occurrence.—
Known from the Wangcun and Wa’ergang sections, where it ranges from the Proconodontus Zone through the Cordylodus angulatus Zone.
Genus Tujiagnathus new genus
Type species.—
Tujiagnathus gracilis n. gen. n. sp.
Diagnosis.—
Slender, cylindrical coniform elements with a minute hook-like cusps. The posterior side is ornamented by numerous tiny nodes. Basal cavity is very deep.
Etymology.—
Tujia, ethnic minority group in western Hunan, South China; gnathus, Greek, jaw.
Tujiagnathus gracilis new genus new species

Figure 25 (1, 2) Prosagittodontus cf. eureka (Müller, Reference Müller1959); (1) Bitiao Formation, Wangcun section, GMPKU2913, posterior view; (2) Shenjiawan Formation, Wa’ergang section, GMPKU2914, posterior view; (3, 6, 7) Serratocambria minuta Müller and Hinz, Reference Müller and Hinz1991; (3) Bitiao Formation, Wangcun section, GMPKU2915, posterior view; (6) Bitiao Formation, Wangcun section, broken specimen, GMPKU2916, posterior view; (7) Bitiao Formation, Wangcun section, GMPKU2917, anterior view; (4, 5, 8–15, 17–22) Teridontus nakamurai (Nogami, Reference Nogami1967); (4) Shenjiawan Formation, Wangcun section, GMPKU2918, lateral view; (5) Shenjiawan Formation, Wangcun section, GMPKU2919, lateral view; (8) Shenjiawan Formation, Wangcun section, GMPKU2920, lateral view; (9) Shenjiawan Formation, Wangcun section, GMPKU2921, lateral view; (10) Shenjiawan Formation, Wangcun section, GMPKU2922, lateral view; (11) Shenjiawan Formation, Wangcun section, GMPKU2923, lateral view; (12) Shenjiawan Formation, Wa’ergang section, GMPKU2924, lateral view; (13) Shenjiawan Formation, Wa’ergang section, GMPKU2925, lateral view; (14) Shenjiawan Formation, Wa’ergang section, GMPKU2926, lateral view; (15) Shenjiawan Formation, Wa’ergang section, GMPKU2927, lateral view; (17) Shenjiawan Formation, Wa’ergang section, GMPKU2928, lateral view; (18) Shenjiawan Formation, Wa’ergang section, GMPKU2929, lateral view; (19) Shenjiawan Formation, Wa’ergang section, GMPKU2930, lateral view; (20) Shenjiawan Formation, Wa’ergang section, GMPKU2931, lateral view; (21) Shenjiawan Formation, Wa’ergang section, GMPKU2932, lateral view; (22) Shenjiawan Formation, Wa’ergang section, GMPKU2933, lateral view; (16, 23) Tujiagnathus gracilis n. gen. n. sp.; (16) Bitiao Formation, Wangcun section, GMPKU2934, holotype, lateral view; (23) Bitiao Formation, Wangcun section, GMPKU2935, lateral view; (24–28) Wangcunella conicus n. gen. n. sp.; (24) Bitiao Formation, Wangcun section, GMPKU2936, lateral view; (25) Bitiao Formation, Wangcun section, GMPKU2937, lateral view; (26) Bitiao Formation, Wangcun section, GMPKU2938, lateral view; (27) Bitiao Formation, Wa’ergang section, GMPKU2939, lateral view; (28) Bitiao Formation, Wa’ergang section, GMPKU2940, holotype, lateral view. Scale bar represents 130 μm (1–28).
Diagnosis.—
As for the genus.
Description.—
Coniform elements, moderate to small, and symmetrical. The cusp is small and hook-like. The base is slender and cylindrical. Both anterior and posterior sides are rounded. The posterior side is ornamented by numerous tiny nodes. Basal cavity is very deep, with the apex extending above the point of flexure of the element.
Etymology.—
gracilis, Latin, gracile or slender.
Types.—
Holotype: GMPKU2934 from the Westergaardodina cf. calix-Prooneotodus rotundatus Zone, Bitiao Formation, Wangcun section, Wangcun village, Yongshun county, Hunan Province.
Materials.—
Nine specimens.
Occurrence.—
Known from Wangcun section, where it occurs in the Westergaardodina cf. calix-Prooneotodus rotundatus Zone.
Comparison.—
In gross morphology, the present species looks like Prooneotodus terashimai, but Tujiagnathus gracilis has conspicuous surface ornamentation, so it is a euconodont (Bengtson, Reference Bengtson1976), whereas P. terashimai is a paraconodont.
Genus Wangcunella new genus
Type species.—
Wangcunella conicus n. gen. n. sp.
Diagnosis.—
Coniform elements without differentiation between cusp and base. Basal cavity is shallow. Cross section is circular.
Etymology.—
Named for the Wangcun section in Hunan Province.
Wangcunella conicus new genus new species
Diagnosis.—
As for the genus.
Description.—
Elements with conical gross morphology. There is no differentiation between cusp and base. The large base tapers rapidly toward the tip. Basal cavity is shallow. Cross section is circular from the apex through the base.
Etymology.—
conicus, Latin, conical, referring to the gross morphology of the element.
Types.—
Holotype: GMPKU2936, from the Westergaardodina cf. calix-Prooneotodus rotundatus Zone, Bitiao Formation, Wangcun section, Wangcun village, Yongshun county, Hunan Province.
Materials.—
18 specimens.
Occurrence.—
Known from the Wangcun and Wa’ergang sections, where it ranges from the Westergaardodina lui-Westergaardodina ani Zone through the Eoconodontus Zone.
Paraconodonts
Class Paraconodontida Müller, Reference Müller1962
[nomen translatum (pro Order Paraconodontida Müller, Reference Müller1962), emended Landing, Reference Landing1995]
Genus Furnishina Müller, Reference Müller1959
Type species.—
Furnishina furnishi Müller, Reference Müller1959.
Furnishina cf. alata Szaniawski, Reference Szaniawski1971

Figure 26 (1–3) Westergaardodina sola n. sp.; (1) Chefu Formation, Wangcun section, GMPKU3031, posterior view; (2) Chefu Formation, Wangcun section, GMPKU3032, posterior view; (3) Chefu Formation, Paibi section, holotype, GMPKU3033, posterior view; (4–9) Westergaardodina tetragonia Dong, Reference Dong1993; (4) Chefu Formation, Wangcun section, GMPKU3034, posterior view; (5) Chefu Formation, Wangcun section, GMPKU3035, posterior view; (6) Chefu Formation, Paibi section, GMPKU3036, posterior view; (7) Chefu Formation, Paibi section, GMPKU3037, posterior view; (8) Chefu Formation, Paibi section, GMPKU3038, posterior view; (9) Chefu Formation, Paibi section, GMPKU3039, posterior view; (10–24) Westergaardodina tricuspidata Müller, Reference Müller1959; (10) Bitiao Formation, Wangcun section, GMPKU3040, posterior view; (11) Bitiao Formation, Wangcun section, GMPKU3041, posterior view; (12) Bitiao Formation, Wangcun section, GMPKU3042, posterior view; (13) Bitiao Formation, Wangcun section, GMPKU3043, posterior view; (14) Bitiao Formation, Wangcun section, GMPKU3044, posterior view; (15) Bitiao Formation, Wangcun section, GMPKU3045, posterior view; (16) Bitiao Formation, Wangcun section, GMPKU3046, posterior view; (17) Bitiao Formation, Wangcun section, GMPKU3047, posterior view; (18) Bitiao Formation, Wangcun section, GMPKU3048, posterior view; (19) Bitiao Formation, Wangcun section, GMPKU3049, posterior view; (20) Bitiao Formation, Wangcun section, GMPKU3050, posterior view; (21) Bitiao Formation, Wangcun section, GMPKU3051, posterior view; (22) Bitiao Formation, Wangcun section, GMPKU3052, posterior view; (23) Bitiao Formation, Wangcun section, GMPKU3053, posterior view; (24) Bitiao Formation, Wangcun section, GMPKU3054, posterior view; (25) Westergaardodina wimani Szaniawski, Reference Szaniawski1971, Bitiao Formation, Wangcun section, GMPKU2200, posterior view; (26) Westergaardodina sp. A, Bitiao Formation, Wangcun section, GMPKU3055, posterior view; (27) Westergaardodina sp. B, Bitiao Formation, Wa’ergang section, GMPKU3056, posterior view; (28) Yongshunella polymorpha Dong and Bergström, 2001a, Huaqiao Formation, Wangcun section, GMPKU3057, cluster; (29, 30) Furnishina cf. alata Szaniawski, Reference Szaniawski1971; (29) Huaqiao Formation, Paibi section, GMPKU3058, posterior view; (30) Huaqiao Formation, Paibi section, GMPKU3059, posterior view; (31–33) Furnishina pernica An, Reference An1982; (31) Huaqiao Formation, Paibi section, GMPKU3060, lateral view; (32) Chefu Formation, Paibi section, GMPKU3061, lateral view; (33) Huaqiao Formation, Paibi section, GMPKU3062, posterior view; (34–36) Hunanognathus tricuspidatus Dong, Reference Dong1993; (34) Huaqiao Formation, Paibi section, GMPKU3063, posterior view; (35) Huaqiao Formation, Paibi section, GMPKU3064, posterior view; (36) Huaqiao Formation, Paibi section, GMPKU3065, posterior view; (37, 38) Shandongodus priscus An, Reference An1982; (37) Huaqiao Formation, Paibi section, GMPKU3066, posterolateral view; (38) Huaqiao Formation, Paibi section, GMPKU3067, lateral view. Relative scale bar represents 183 μm (1, 2, 5, 10–24, 28, 34), 323 μm (3), 172 μm (4, 6, 7, 9, 25, 29, 32, 33, 35, 36), 238 μm (8), 373 μm (26), 472 μm (27), 157 μm (30, 31), 139 μm (37, 38).
cf.1971 Furnishina alata Reference SzaniawskiSzaniawski, p. 406, pl. 1, figs. 3, 4, pl. 3, figs. 3–5.
1987Furnishina aff. furnishi Müller; Reference AnAn, pl. 3, fig. 8.
1991Furnishina cf. alata Szaniawski; Reference Müller and HinzMüller and Hinz, p. 16, pl. 7, figs. 18, 20.
1993Furnishina cf. alata Szaniawski; Reference DongDong, p. 350, pl. 2, figs. 11, 13, 15.
2001aFurnishina cf. alata Szaniawski; Dong and Bergström, p. 960, pl. 1, figs. 1, 3, 10.
2006Furnishina cf. F. alata Szaniawski; Reference Qi, Bagnoli and WangQi et al., p. 182, pl. 2, fig. 15.
2008Furnishina cf. alata Szaniawski; Reference Bagnoli, Qi and WangBagnoli et al., p. 112, fig. 3 (A).
Materials.—
48 specimens.
Occurrence.—
Present in the Paibi and Wangcun sections from the Shandongodus priscus-Hunanognathus tricuspidatus Zone through the Westergaardodina matsushita-Westergaardodina grandidens Zone.
Remarks.—
The specimens at hand agree in gross morphology with Furnishina alata but differ in the posterior keel, which is not developed or present only along the cusp.
Furnishina bigeminata Dong, Reference Dong1993
1993Furnishina bigeminata Reference DongDong, p. 350, pl. 4, figs. 3, 4, 6, 14.
2001aFurnishina bigeminata Dong; Reference Dong and BergströmDong and Bergström, p. 960, pl. 3, fig. 17, pl. 5, fig. 14.
2006Furnishina bigeminata Dong; Reference Qi, Bagnoli and WangQi et al., p. 186, pl. 2, fig. 6.
2008Furnishina bigeminata Dong; Reference Bagnoli, Qi and WangBagnoli et al., p. 112, fig. 3 (B).
Materials.—
688 specimens.
Occurrence.—
Present in the Paibi, Wangcun, and Wa’ergang sections, where it ranges from the Gapparodus bisulcatus-Westergaardodina brevidens Zone through the Cordylodus proavus Zone.
Remarks.—
The present specimens agree well with Furnishina bigeminata as described and figured by Dong and Bergström (Reference Dong and Bergström2001a).
Furnishina dayangchaensis Chen and Gong, Reference Chen and Gong1986
1986Furnishina dayangchaensis Reference Chen and GongChen and Gong, p. 144, pl. 17, figs. 2, 3, 6, 8, 9, 14, pl. 19, fig. 11, text-fig. 50 (2).
2000Furnishina furnishi Müller; Reference Pyle and BarnesDubinina, p. 168, pl. 2, fig. 17. (part)
Materials.—
78 specimens.
Occurrence.—
Present in the Wangcun and Wa’ergang sections, where it ranges from the Westergaardodina lui-Westergaardodina ani Zone through the Cordylodus proavus Zone.
Remarks.—
The present specimens agree well with Furnishina dayangchaensis as described and figured by Chen and Gong (Reference Chen and Gong1986).
Furnishina furnishi Müller, Reference Müller1959

Figure 27 (1–9) Furnishina furnishi Müller, Reference Müller1959; (1) Bitiao Formation, Wangcun section, GMPKU2601, posterior view; (2) Bitiao Formation, Wangcun section, GMPKU2602, posterior view; (3) Bitiao Formation, Wangcun section, GMPKU2603, posterior view; (4) Bitiao Formation, Wangcun section, GMPKU2604, posterior view; (5) Bitiao Formation, Wangcun section, GMPKU2605, posterior view; (6) Bitiao Formation, Wangcun section, GMPKU2606, posterior view; (7) Bitiao Formation, Wangcun section, GMPKU2607, posterior view; (8) Bitiao Formation, Wangcun section, GMPKU2608, posterior view; (9) Shenjiawan Formation, Wa’ergang section, GMPKU2609, posterior view; (10–16) Furnishina cf. furnishi Müller, Reference Müller1959; (10) Bitiao Formation, Wangcun section, GMPKU2610, oblique posterior view; (11) Bitiao Formation, Wangcun section, GMPKU2611, lateral view; (12) Bitiao Formation, Wangcun section, GMPKU2612, lateral-posterior view; (13) Bitiao Formation, Wangcun section, GMPKU2613, posterior view; (14) Bitiao Formation, Wangcun section, GMPKU2614, lateral view; (15) Bitiao Formation, Wangcun section, GMPKU2615, posterior view; (16) Bitiao Formation, Wangcun section, GMPKU2616, posterior view; (17) Furnishina gladiata Müller and Hinz, Reference Müller and Hinz1991, Bitiao Formation, Wangcun section, GMPKU2617, oblique posterior view; (18) Furnishina cf. gladiata Müller and Hinz, Reference Müller and Hinz1991, Huaqiao Formation, Wangcun section, GMPKU2618, lateral-posterior view; (19, 20, 22) Furnishina kleithria Müller and Hinz, Reference Müller and Hinz1991; (19) Chefu Formation, Wangcun section, GMPKU2619, posterior view; (20) Chefu Formation, Wangcun section, GMPKU2620, posterior view; (22) Chefu Formation, Wangcun section, GMPKU2621, posterior view; (21) (23–25) Furnishina kranzae Müller and Hinz, Reference Müller and Hinz1991; (21) Bitiao Formation, Wangcun section, GMPKU2622, posterior view; (23) Bitiao Formation, Wangcun section, GMPKU2623, posterior view; (24) Bitiao Formation, Wangcun section, GMPKU2624, posterior view; (25) Bitiao Formation, Wangcun section, GMPKU2625, posterior view. Scale bar represents 132 μm (1–25).
1959Furnishina furnishi Reference MüllerMüller, p. 452, pl. 11, figs. 5, 6, 9, 11–13, 15, text-fig. 6D.
1971Furnishina asymmetrica Müller; Reference MüllerMüller, p. 8, pl. 1, figs. 9, 12, 14–16.
1971Furnishina furnishi Müller; Reference Müller and NogamiMüller and Nogami, p. 14, pl. 1, fig. 5.
1972Furnishina furnishi Müller; Reference Müller and NogamiMüller and Nogami, fig. 1A.
1973Furnishina asymmetrica Müller; Reference MüllerMüller, p. 39, pl. 1, figs. 6, 8.
? 1976Furnishina furnishi Müller; Reference Abaimova and ErgalievAbaimova and Ergaliev, p. 391, pl. 14, figs. 1, 2.
? 1976Furnishina furnishi Müller; Miller et al., p. 595, pl. 1, figs. 8–12.
1979Furnishina furnishi Müller; Reference BednarczykBednarczyk, p. 427, pl. 1, figs. 1–3, 5–9, 12, pl. 3, figs. 14, 20, 21.
1981Furnishina furnishi Müller; Miller in Reference RobisonRobison, p. W112, fig. 64, 7a, b.
1986Furnishina furnishi Müller; Reference Chen, Zhang and YuChen et al., p. 367, pl. 1, fig. 17.
1988Furnishina furnishi Müller; Reference Lee and LeeLee and Lee, pl. 1, figs. 16, 17.
1991Furnishina furnishi Müller; Reference Müller and HinzMüller and Hinz, p. 17, pl. 13, figs. 1–7, 11, 12, text-fig. 8 (A).
? 2000Furnishina furnishi Müller; Reference Göncüoğlu and KozurGöncüoğlu and Kozur, p. 599, fig. 4 (h).
? 2000Furnishina furnishi Müller; Reference Pyle and BarnesDubinina, p. 168, pl. 3, figs. 9–10 (not pl. 1, figs. 8, 9, 26, pl. 3, figs. 2, 3, 12, 23, 25). (part)
2009Furnishina furnishi Müller; Reference Lee, Lee, Miller and JeongLee et al., p. 421, fig. 6 (17).
Materials.—
30 specimens.
Occurrence.—
Known from the Wangcun and Wa’ergang sections, where it ranges from the Westergaardodina lui-Westergaardodina ani Zone through the Cordylodus proavus Zone.
Remarks.—
Müller and Hinz (Reference Müller and Hinz1991) emended the diagnosis of Furnishina furnishi. Previously, it was defined very broadly and included a variety of forms that could not be assigned properly to another species. The new diagnosis restricted the occurrence of F. furnishi, so that it ranges from the trilobite zone 3 through zone 5 in the upper Cambrian of Sweden. Accordingly, many elements previously refered to F. furnishi by different authors, especially Chinese authors, should be excluded from this species based on their characters and occurrences.
Furnishina cf. furnishi Müller, Reference Müller1959
cf.1959 Furnishina furnishi Reference MüllerMüller, p. 452, pl. 11, figs. 5, 6, 9, 11–13, 15, text-fig. 6D.
cf.1991 Furnishina furnishi Müller; Reference Müller and HinzMüller and Hinz, p. 17, pl. 13, figs. 1–7, 11, 12; text-fig. 8 (A).
2000Furnishina furnishi Müller; Reference Pyle and BarnesDubinina, p. 168, pl. 3, fig. 11. (part)
2007Furnishina furnishi Müller; Reference Landing, Westrop and KeppieLanding et al., p. 922, fig. 9 (c).
2009Furnishina furnishi Müller; Reference Lee, Lee, Miller and JeongLee et al., p. 423, fig. 7 (16), p. 425, fig. 8 (10).
Materials.—
16 specimens.
Occurrence.—
Known from the Wangcun and Wa’ergang sections, where it ranges from the Westergaardodina lui-Westergaardodina ani Zone through the Eoconodontus Zone.
Remarks.—
Similar to Furnishina furnishi but differs in the posterior side, which is not convexly domed, and the median keel is indistinct.
Furnishina gladiata Müller and Hinz, Reference Müller and Hinz1991
1959Scandodus n. sp. b Reference MüllerMüller, p. 464, pl. 12, fig. 5.
1991Furnishina gladiata Reference Müller and HinzMüller and Hinz, p. 29, pl. 16, figs. 1–13, 15, text-fig. 8 (S).
Materials.—
Three specimens.
Occurrence.—
Known from the Wangcun sections, where it occurs in the Westergaardodina lui-Westergaardodina ani Zone.
Remarks.—
The specimens at hand agree in all respects with Müller and Hinz’s (Reference Müller and Hinz1991) species.
Furnishina cf. gladiata Müller and Hinz, Reference Müller and Hinz1991
cf. 1959Scandodus n. sp. b Reference MüllerMüller, p. 464, pl. 12, fig. 5.
cf. 1991Furnishina gladiata Reference Müller and HinzMüller and Hinz, p. 29, pl. 16, figs. 1–13, 15, text-fig. 8 (S).
Materials.—
Two specimens.
Occurrence.—
Known from the Wangcun section, where it ranges from Shandongodus priscus-Hunanognathus tricuspidatus Zone through the Westergaardodina cf. calix-Prooneotodus rotundatus Zone.
Remarks.—
Similar to Furnishina gladiata, but it differs in its anterolateral costae that do not appear as large laminae.
Furnishina kleithria Müller and Hinz, Reference Müller and Hinz1991
1991Furnishina kleithria Reference Müller and HinzMüller and Hinz, p. 21, pl. 15, figs. 1–5, 7–10, 16, text-fig. 8 (P, Q).
2006Furnishina kleithria Müller and Hinz; Reference Qi, Bagnoli and WangQi et al., p. 186, pl. 2, fig. 7.
2008Furnishina kleithria Müller and Hinz; Reference Bagnoli, Qi and WangBagnoli et al., p. 112, fig. 3 (R).
Materials.—
10 specimens.
Occurrence.—
Known from the Wangcun section, where it ranges from the Gapparodus bisulcatus-Westergaardodina brevidens Zone through the Westergaardodina cf. calix-Prooneotodus rotundatus Zone.
Remarks.—
The present specimens at hand agree with Furnishina kleithria as described and illustrated by Müller and Hinz (Reference Müller and Hinz1991).
Furnishina kranzae Müller and Hinz, Reference Müller and Hinz1991
1991Furnishina kranzae Reference Müller and HinzMüller and Hinz, pl. 12, figs. 1, 2, 6, 8, 12–14, 18, fig. 8 (B).
Materials.—
Nine specimens.
Occurrence.—
Known from the Paibi and Wangcun sections, where it ranges from Westergaardodina lui-Westergaardodina ani Zone Zone through the Westergaardodina cf. calix-Prooneotodus rotundatus Zone.
Remarks.—
The specimens at hand agree with Furnishina kranzae as described and figured by Müller and Hinz (Reference Müller and Hinz1991).
Furnishina cf. kranzae Müller and Hinz, Reference Müller and Hinz1991

Figure 28 (1) Furnishina cf. kranzae Müller and Hinz, Reference Müller and Hinz1991, Bitiao Formation, Wangcun section, GMPKU2626, posterior view; (2, 3) Furnishina longibasis Bednarczyk, Reference Bednarczyk1979; (2) Bitiao Formation, Wangcun section, GMPKU2627, lateral view; (3) Bitiao Formation, Wangcun section, GMPKU2628, lateral-anterior view; (4, 5) Furnishina cf. ovata Müller and Hinz, Reference Müller and Hinz1991; (4) Bitiao Formation, Wangcun section, GMPKU2629, posterior view; (5) Bitiao Formation, Wangcun section, GMPKU2630, posterior view; (6–10, 13) Furnishina primitiva Müller, Reference Müller1959; (6) Bitiao Formation, Wangcun section, GMPKU2631, posterior view; (7), Shenjiawan Formation, Wa’ergang section, GMPKU2632, posterior view; (8) Shenjiawan Formation, Wa’ergang section, GMPKU2633, posterior view; (9) Shenjiawan Formation, Wa’ergang section, GMPKU2634, posterior view; (10) Shenjiawan Formation, Wa’ergang section, GMPKU2635, posterior view; (13) Shenjiawan Formation, Wa’ergang section, GMPKU2636, posterior view; (11, 14–17) Furnishina cf. quadrata Müller, Reference Müller1959; (11) Bitiao Formation, Wangcun section, GMPKU2637, oblique posterior view; (14) Chefu Formation, Wangcun section, GMPKU2638, posterior view; (15) Bitiao Formation, Wangcun section, GMPKU2639, posterior view; (16) Bitiao Formation, Wangcun section, GMPKU2640, posterior view; (17) Bitiao Formation, Wangcun section, GMPKU2641, top view; (12) Furnishina quadrata Müller, Reference Müller1959, Bitiao Formation, Wangcun section, GMPKU2642, top view; (18) Furnishina tortilis (Müller, Reference Müller1959) Bitiao Formation, Wangcun section, GMPKU2643, posterior view; (19–23) Furnishina wangcunensis n. sp.; (19) Bitiao Formation, Wangcun section, GMPKU2644, lateral view; (20) Bitiao Formation, Wangcun section, GMPKU2645, lateral view; (21) Bitiao Formation, Wangcun section, GMPKU2646, lateral view; (22) Bitiao Formation, Wangcun section, holotype, GMPKU2647, lateral view; (23) Bitiao Formation, Wangcun section, GMPKU2648, posterior view; (24–26) Gapparodus bisulcatus (Müller, Reference Müller1959); (24) Huaqiao Formation, Wangcun section, GMPKU2649, lateral view; (25) Huaqiao Formation, Wangcun section, GMPKU2650, lateral view; (26) Huaqiao Formation, Wangcun section, GMPKU2651, lateral view. Relative scale bar represents 154 μm (1–26).
cf.1991 Furnishina kranzae Reference Müller and HinzMüller and Hinz, pl. 12, figs. 1, 2, 6, 8, 12–14, 18, fig. 8 (B).
1993Furnishina cf. kranzae Müller and Hinz; Reference DongDong, p. 350, pl. 4, figs. 1, 2, 5.
2001aFurnishina cf. kranzae Müller and Hinz; Reference Dong and BergströmDong and Bergström, p. 961, pl. 5, fig. 9.
Materials.—
18 specimens.
Occurrence.—
Known from the Paibi, Wangcun and Wa’ergang sections, where it ranges from the Westergaardodina quadrata Zone through the Westergaardodina cf. calix-Prooneotodus rotundatus Zone.
Remarks.—
Differs from the typical Furnishina kranzae in having indistinctly laterally deflected cusps and in the weakly developed posterior keels. Our specimens also occur in stratigraphically older strata than F. kranzae.
Furnishina longibasis Bednarczyk, Reference Bednarczyk1979
1979Furnishina longibasis Reference BednarczykBednarczyk, p. 427, pl. 1, figs. 1, 4.
1991Furnishina longibasis Bednarczyk; Reference Müller and HinzMüller and Hinz, p. 21, pl. 11, figs. 10, 12.
2006Furnishina longibasis Bednarczyk; Reference Qi, Bagnoli and WangQi et al., p. 186, pl. 2, figs. 2, 4 (not 1, 3). (part)
2009Furnishina longibasis Bednarczyk; Reference Lee, Lee, Miller and JeongLee et al., p. 420, fig. 6 (3).
Materials.—
Six specimens.
Occurrence.—
Known from the Wangcun section, where it ranges from the Shandongodus priscus-Hunanognathus tricuspidatus Zone through the Westergaardodina cf. calix-Prooneotodus rotundatus Zone.
Remarks.—
The specimens at hand agree in all respects with Furnishina longibasis.
Furnishina cf. ovata Müller and Hinz, Reference Müller and Hinz1991
cf.1991 Furnishina ovata Reference Müller and HinzMüller and Hinz, p. 22, pl. 8, figs. 7, 8, 10–22, text-fig. 8 (T).
Materials.—
Four specimens.
Occurrence.—
Known from the Wangcun and Wa’ergang sections, where it ranges from Westergaardodina lui-Westergaardodina ani Zone through the Eoconodontus Zone.
Remarks.—
Similar to Furnishina ovata but differs in its anterolateral costae, which are not restricted to the base but extend to the cusp and are more pronounced.
Furnishina pernica An, Reference An1982
1982Furnishina pernica Reference AnAn, p. 133, pl. 4, figs. 1–4, pl. 8, fig. 4.
1993Furnishina pernica An; Reference DongDong, pl. 5, figs. 4, 5, 9.
2001aFurnishina pernica An; Reference Dong and BergströmDong and Bergström, p. 961, pl. 5, figs. 1, 13.
? 2009 Furnishina pernica An; Reference Lee, Lee, Miller and JeongLee et al., p. 425, fig. 8 (4).
Materials.—
26 specimens.
Occurrence.—
Known from the Paibi and Wangcun sections, where it ranges from the Shandongodus priscus-Hunanognathus tricuspidatus Zone through the Westergaardodina quadrata Zone.
Remarks.—
The specimens at hand agree well with Furnishina pernica.
Furnishina primitiva Müller, Reference Müller1959
1959Furnishina primitiva Reference MüllerMüller, p. 453, pl. 11, figs. 1 (?), 2–4.
1971Furnishina primitiva Müller; Reference MüllerMüller, pl. 1, fig. 11.
1976Furnishina primitiva Müller; Reference Abaimova and ErgalievAbaimova and Ergaliev, pl. 14, fig. 8.
1986Furnishina primitiva Müller; Reference Chen and GongChen and Gong, p. 146, pl. 17, figs. 7, 11, 18, pl. 18, fig. 11 (not pl. 20, fig. 8), text-fig. 51 (1–3, not 4). (part)
1987Furnishina primitiva Müller; Reference AnAn, p. 106, pl. 3, fig. 10.
1988Furnishina primitiva Müller; Reference Lee and LeeLee and Lee, pl. 1, figs. 14, 15.
1991Furnishina primitiva Müller; Reference Müller and HinzMüller and Hinz, p. 23, pl. 12, figs. 3, 4, 7, 9, 10, 15–17, 19, text-fig. 8 (C, D).
non.1993 Furnishina primitiva Müller; Reference LandingLanding, p. 6, fig. 4 (3).
2009Furnishina primitiva Müller; Reference Lee, Lee, Miller and JeongLee et al., p. 421, fig. 6 (2, 4, not 7). (part)
Materials.—
18 specimens.
Occurrence.—
Known from the Wangcun and Wa’ergang sections, where it ranges from Westergaardodina lui-Westergaardodina ani Zone through the Cordylodus intermedius Zone.
Remarks.—
The specimens at hand agree in all respects with Müller’s species.
Furnishina quadrata Müller, Reference Müller1959
1959Furnishina quadrata Reference MüllerMüller, p. 453, pl. 12, figs. 2, 4, 9, fig. 6C.
1966Furnishina quadrata Müller; Reference NogamiNogami, p. 355, pl. 9, figs. 3, 4.
1976Furnishina quadrata Müller; Reference Abaimova and ErgalievAbaimova and Ergaliev, p. 392, pl. 14, figs. 3, 4, 6, 7.
1979Furnishina quadrata Müller; Reference BednarczykBednarczyk, p. 428, pl. 1, figs. 15, 16.
1981Furnishina quadrata Müller; Reference Miller, Sundberg, Harma and WrightMiller et al., fig. 4 (E).
? 1985 Furnishina quadrata Müller; Reference WangWang, p. 226, pl. 3, fig. 14, pl. 8, fig. 14.
? 1986Furnishina quadrata Müller; Reference Chen and GongChen and Gong, p. 147, pl. 17, fig. 17, text-fig. 52 (2).
1991Furnishina quadrata Müller; Reference Müller and HinzMüller and Hinz, p. 23, pl. 9, figs. 1–13.
2001aFurnishina quadrata Müller; Reference Dong and BergströmDong and Bergström, p. 961, pl. 3, fig. 16.
? 2006Furnishina quadrata Müller; Reference Qi, Bagnoli and WangQi et al., p. 185, pl. 1, figs. 13–14.
Materials.—
Five specimens.
Occurrence.—
Known only from the Wangcun section, where it occurs from the Westergaardodina quadrata Zone through the Westergaardodina lui-Westergaardodina ani Zone.
Remarks.—
The specimens at hand agree closely with Müller’s description.
Furnishina cf. quadrata Müller, Reference Müller1959
cf.1959 Furnishina quadrata Reference MüllerMüller, pl. 12, figs. 2, 4, 9, fig. 6C.
cf.1991 Furnishina quadrata Müller; Reference Müller and HinzMüller and Hinz, pl. 9, figs. 1–13.
1993Furnishina cf. quadrata Müller; Reference DongDong, p. 351, pl. 5, figs. 7, 8, 10, 18.
2001aFurnishina cf. quadrata Müller; Dong and Bergström, p. 962, pl. 3, fig. 13.
Materials.—
36 specimens.
Occurrence.—
Known from the Paibi and Wangcun sections, where it is present in the Shandongodus priscus-Hunanognathus tricuspidatus Zone through the Westergaardodina cf. calix-Prooneotodus rotundatus Zone.
Remarks.—
Differs from the type specimen of Furnishina quadrata in that the anterior sides of the element are not concave but slightly convex. Its first appearance also occurs stratigraphically lower than typical F. quadrata.
Furnishina tortilis (Müller, Reference Müller1959)
1959Scandodus tortilis Reference MüllerMüller, p. 464, pl. 12, figs. 7, 8, 10, text-fig. 9.
1971Scandodus tortilis Müller; Reference MüllerMüller, pl. 2, fig. 1.
1979Scandodus tortilis Müller; Reference BednarczykBednarczyk, p. 434, pl. 4, fig. 15.
1981Proscandodus tortilis (Müller); Miller in Reference RobisonRobison, p. 113, fig. 64 (5).
1991Furnishina tortilis (Müller); Reference Müller and HinzMüller and Hinz, p. 24, pl. 14, figs. 2–19, text-fig. 8 (N, O).
? 2006Furnishina tortilis (Müller); Reference Qi, Bagnoli and WangQi et al., p. 185, pl. 1, figs. 5–9.
2009Furnishina tortilis (Müller); Reference Lee, Lee, Miller and JeongLee et al., p. 425, fig. 8 (1, ?13).
Materials.—
Three specimens.
Occurrence.—
Known only from the Wangcun section, where it occurs in the Westergaardodina lui-Westergaardodina ani Zone.
Remarks.—
The specimens at hand agree well with Furnishina tortilis.
Furnishina wangcunensis new species
Diagnosis.—
Slender coniform elements. Cusp is distinctively recurved. The posterior costa and two anterolateral costae are sharp. Basal cavity is very deep.
Description.—
Slender coniform elements with the cusp distinctively recurved toward the posterior side. Elements usually subsymmetrical but rarely asymmetrical due to lateral bending. Anterior side is convex, whereas the two posterolateral sides are concave. The posterior costa and two anterolateral costae are sharp, extending from the base up to a point that is above the recurved part of the cusp. The cross section is rounded apically and passes into subtriangular along the rest of the entire element, except for the base, where the cross section is subtriangular or irregularly rectangular. The basal cavity is very deep, with the apex above the recurved point of cusp.
Etymology.—
Named for the Wangcun section in Hunan Province.
Types.—
Holotype: GMPKU2647, from the Westergaardodina cf. calix-Prooneotodus rotundatus Zone, Furongian (upper Cambrian), Bitiao Formation, Wangcun section, Wangcun village, Yongshun County, Hunan Province.
Materials.—
98 specimens.
Occurrence.—
Known from the Wangcun section where it is present in the Westergaardodina cf. calix-Prooneotodus rotundatus Zone.
Comparison.—
Furnishina wangcunensis n. sp. differs from Furnishina dayangchaensis in its more slender elements, distinctively recurved cusps, and sharp posterior costae and anterolateral costae.
Genus Hunanognathus Dong, Reference Dong1993
Type species.—
Hunanognathus tricuspidatus Dong, Reference Dong1993.
Hunanognathus tricuspidatus Dong, Reference Dong1993
Figures 20.9, 20.10, 26.34–26.36
? 1987Westergaardodina amplicava (Müller); Reference AnAn, pl. 3, fig. 14.
1993Hunanognathus tricuspidatus Reference DongDong, p. 352, pl. 3, figs. 1–3.
2001aHunanognathus tricuspidatus Dong; Reference Dong and BergströmDong and Bergström, p. 965, pl. 4, figs. 2–4.
Materials.—
60 specimens.
Occurrence.—
Known from the Paibi and Wangcun sections, where it ranges from the Shandongodus priscus-Hunanognathus tricuspidatus Zone through the Westergaardodina quadrata Zone.
Remarks.—
The specimens at hand agree well with Hunanognathus tricuspidatus described and figured by Dong and Bergström (2001a). A single specimen referred to as Furnishina? sp. indet. by Müller and Hinz (Reference Müller and Hinz1991, p. 25, pl. 14, fig. 1) was found in Zone 1 at Gum, Sweden, and a similar specimen was recovered in the present study in the lower part of the Westergaardodina quadrata Zone of the Chefu Formation. In gross mophology, both these specimens are simliar to H. tricuspidatus, but they differ from the latter in having only one small denticle. It is probable that these specimens are closely related to H. tricuspidatus, but additional specimens are needed to assess this possible relationship.
Genus Laiwugnathus An, Reference An1982
Diagnosis.—
Coniform elements, triangular in outline. The anterior side is convex, whereas the posterior side is concave, with a prominent median keel. Both sides are fused along the upward arched basal margin.
Description.—
Coniform elements, triangular in outline, without a clear boundary between cusp and base. Anterior side is slightly convex. Posterior side is concave and is characterized by a slender median keel ending with an opening. The keel tapers gradually upward. Both anterior and posterior sides are fused along the basal margin. No basal cavity could be observed. Lateral edge of element is straight and sharp. Basal margin is sharp and is arched upward. The end of the keel is higher than the highest part of the arched basal margin.
Etymology.—
Named for its provenance in the Chinese Province of Hunan.
Types.—
Holotype: GMPKU2722, from Gapparodus bisulcatus-Westergaardodina brevidens Zone, middle Cambrian, Huaqiao Formation, Wangcun section, Wangcun village, Yongshun County, Hunan Province.
Materials.—
Nine specimens.
Occurrence.—
Present in the Wangcun section, where it occurs in the Gapparodus bisulcatus-Westergaardodina brevidens Zone.
Comparison.—
Laiwugnathus hunanensis n. sp. differs from Laiwugnathus fenghuangesis An, Reference An1982 in its much more slender median keel and the absence of a basal cavity. It differs from Laiwugnathus kouzhenensis An, Reference An1982 in its much shorter median keel and upward-arched basal margin. The median keel of L. kouzhenensis is much longer, ending with an opening shaped like a loudspeaker and extending below the basal margin. L. hunanensis differs from Laiwugnathus graoilentus An and Mei, Reference An and Mei1994 in that the keel of the latter is more slender and solid.
Remarks.—
Specimens of various species of Laiwugnathus are very common in North China but are relatively rare in Hunan.
Laiwugnathus cf. kouzhenensis An, Reference An1982
1982Laiwugnathus cf. kouzhenensis An, Reference An1982, p. 135, pl. 2, figs. 12–13.
Materials.—
Five specimens.
Occurrence.—
Present in the Wangcun section, where it occurs in the Westergaardodina matsushitai-Westergaardodina grandidens Zone.
Remarks.—
Similar to Laiwugnathus kouzhenensis, but the specimens at hand differ in the longitudinal keel. In L. kouzhenensis, the median keel is prominent, ending with an opening shaped like a loudspeaker. In the present specimens, the boundary between the median keel and the posterior side of the upper part of the cusp is not distinct.
Laiwugnathus transitans new species
2001aWestergaardodina sp. Reference Dong and BergströmDong and Bergström, p. 963, pl. 1, fig. 21.
Diagnosis.—
Coniform elements with a large cusp and small base. The anterior side is convex, whereas the posterior side is concave, with a prominent median rounded keel. Both sides are fused along the straight basal margin.
Description.—
Coniform elements with a long, narrow cusp. The small base extends laterally, like a very short lateral projection typical of Westergaardodina. Anterior side is slightly convex. Posterior side is concave and is characterized by a rounded, median keel that is long and large that ends with cone shaped opening. Both anterior and posterior sides are fused along the basal margin. No basal cavity could be observed. Lateral edge of element is straight and sharp. Basal margin is sharp and almost straight. The end of the keel is near the basal margin.
Etymology.—
Latin, transitans, transitional.
Types.—
Holotype: GMPKU2726, from Westergaardodina matsushitai-Westergaardodina grandidens Zone, middle Cambrian, Chefu Formation, Wangcun section, Wangcun village, Yongshun County, Hunan Province.
Materials.—
Ten specimens.
Occurrence.—
Present in the Paibi and Wangcun sections, where it ranges from the Westergaardodina matsushitai-Westergaardodina grandidens Zone through the Westergaardodina lui-Westergaardodina ani Zone.
Comparison.—
Laiwugnathus transitans n. sp. differs from Laiwugnathus kouzhenensis in its base which inclines downwards like two short lateral projections of the genus Westergaardodina.
Remarks.—
Laiwugnathus transitans n. sp. is probably the transitional form between Laiwugnathus and tricuspidate Westergaardodina.
Genus Lugnathus new genus
Type species.—
Lugnathus hunanensis n. gen. n. sp.
Diagnosis.—
Large, simple, coniform elements, proclined, asymmetrical and laterally compressed, with a carina on one lateral side. Anterior and posterior edges lack costae. Basal cavity is very deep, the apex of which extends nearly to the tip of the cusp.
Etymology.—
In honor of the late Professor Lu Yan-hao, Nanjing Institute of Geology and Palaeontology, Chinese Academy of Sciences; Acadimician of Chinese Academy of Sciences, the name is composed from Lu and a derivative of Greek, gnathus, jaw.
Remarks.—
In gross morphology, Lugnathus is somewhat similar to Proconodontus but differs from the latter in lacking either an anterior or posterior costa. Also, Lugnathus is a paraconodont, which is verified by our histological investigtion of specimens recovered from Hunan (Fig. 29).

Figure 29 Conodont images using oil immersion techniques (Dong et al., 2005c) with Differential Interference Contrast (Nomarski) illumination, showing pataconodont structure. (1–4) Lugnathus hunanensis n. gen. n. sp.; (1) overview, GMPKU3068; (2) close-up of (1), showing the growth lamellae of paraconodont; (3) close-up of the tip, showing the growth lamellae of paraconodont in detail; (4) close-up of (1) showing the structure of the basal tissue which intergrades with the core of calcosphere (dentine). Relative scale bar represents 46 μm (1), 27 μm (2), 11 μm (3), 6 μm (4).
Lugnathus hunanensis new genus new species
Diagnosis.—
As for the genus.
Description.—
Large, hollow, coniform elements, proclined, asymmetrical, laterally compressed, with carina on one lateral side. The apex is pointed. The cusp is slender, with a pointed tip. Anterior and posterior edges lack costae. The basal cavity is very deep, the apex of which extends nearly to tip of cusp.
Etymology.—
Named for Hunan Province.
Types.—
Holotype: GMPKU2702, from Cordylodus intermedius Zone, Furongian (upper Cambrian), Panjiazui Formation, Wa’ergang section, Wa’ergang village, Taoyuan County, Hunan Province.
Materials.—
120 specimens.
Occurrence.—
Known from the Wa’ergang and Wangcun sections, where it ranges from the Eoconodontus Zone through the Cordylodus lindstromi Zone (upper part).
Genus Miaognathus new genus
Type species.—
Miaognathus multicostatus n. gen. n. sp.
Diagnosis.—
Large coniform elements, proclined and asymmetrical. It is strongly laterally compressed. Basal cavity is very large and deep. The posterior edge has a costa, and the anterior edge is without costae. One to four costae are on each of the lateral sides.
Etymology.—
Miao, ethnic minority group in werstern Hunan, South China; gnathus, Greek, jaw.
Miaognathus multicostatus new genus new species
Diagnosis.—
As for the genus.
Description.—
Large-sized coniform, proclined and asymmetrical. It is strongly laterally compressed. Basal cavity is very large and deep, the apex of which extends nearly to tip. The posterior edge is characterized by a sharp costa, whereas the anterior edge is without costa. One to four costae are on each of the lateral sides. Cross-section is sharp subtriangular except for the apex.
Etymology.—
multicostatus, Latin, referring to the multiple costae.
Types.—
Holotype: GMPKU2733 from the Eoconodontus Zone, Furongian (upper Cambrian), Shenjiawan Formation, Wa’ergang section, Wa’ergang village, Taoyuan County, Hunan Province.
Materials.—
20 specimens.
Occurrence.—
Known from the Wa’ergang section, where it occurs in the Eoconodontus Zone.
Comparison.—
Miaognathus multicostatus n. gen. n. sp. is more or less similar to species of Coelocerodontus in gross morphology, but it differs from the latter in its much larger size, lack of an anterior keel, and the presence of more than one costa on its lateral sides. Also, their stratigraphic ranges are different.
Remarks.—
The species of Coelocerodontus are euconodonts, whereas Miaognathus multicostatus n. gen. n. sp. is probably a paraconodont.
Genus Muellerodus Miller, Reference Miller1980
Type species.—
Distacodus (?) cambricus Müller, Reference Müller1959.
Remarks.—
The name Muellerodus Miller, Reference Miller1980 was proposed to replace Muellerina Szaniawski, Reference Szaniawski1971, which is a homonym of an ostracode genus proposed in 1965.
Muellerodus cambricus (Müller, Reference Müller1959)

Figure 30 (1–3, 6) Muellerodus cambricus (Müller, Reference Müller1959); (1) Shenjiawan Formation, Wa’ergang section, GMPKU2744, lateral view; (2) Shenjiawan Formation, Wa’ergang section, GMPKU2745, lateral view; (3) Shenjiawan Formation, Wa’ergang section, GMPKU2746, lateral view; (6) Shenjiawan Formation, Wa’ergang section, GMPKU2747, lateral view; (4) Muellerodus guttulus Müller and Hinz, Reference Müller and Hinz1991, Bitiao Formation, Wangcun section, GMPKU2748, posterior view; (5, 11, 12) Muellerodus? obliquus (An, Reference An1982); (5) Bitiao Formation, Wangcun section, GMPKU2749, lateral view; (11) Bitiao Formation, Wangcun section, GMPKU2750, lateral view; (12) Huaqiao Formation, Paibi section, GMPKU2751, lateral view; (7–10) Muellerodus? oelandicus (Müller, Reference Müller1959); (7) Huaqiao Formation, Wangcun section, GMPKU2752, lateral view; (8) Bitiao Formation, Wangcun section, GMPKU2753, lateral view; (9) Bitiao Formation, Wangcun section, GMPKU2754, lateral view; (10) Shenjiawan Formation, Wangcun section, GMPKU2755, lateral view; (13–21) Muellerodus pomeranensis (Szaniawski, Reference Szaniawski1971); (13) Bitiao Formation, Wangcun section, GMPKU2756, posterior view; (14) Bitiao Formation, Wangcun section, GMPKU2757, posterolateral view; (15) Bitiao Formation, Wangcun section, GMPKU2758, posterior view; (16) Bitiao Formation, Wangcun section, GMPKU2759, posterior view; (17) Bitiao Formation, Wangcun section, GMPKU2760, posterior view; (18) Bitiao Formation, Wangcun section, GMPKU2761, oblique posterior view; (19) Bitiao Formation, Wangcun section, GMPKU2762, posterior view; (20) Bitiao Formation, Wangcun section, GMPKU2763, posterior view; (21) Bitiao Formation, Wangcun section, GMPKU2764, posterior view; (22–25) Nogamiconus sinensis (Nogami, Reference Nogami1966); (22) Huaqiao Formation, Wangcun section, GMPKU2765, posterior view; (23) Chefu Formation, Wangcun section, GMPKU2766, posterior view; (24) Chefu Formation, Wangcun section, GMPKU2767, posterior view; (25) Chefu Formation, Wangcun section, GMPKU2768, posterior view. Relative scale bar represents 123 μm (1, 2, 4, 6, 8, 11, 16, 17, 22, 25), 117 μm (3, 5, 7, 9, 10, 12–15, 18–21), 159 μm (23, 24).
1959Distacodus (?) cambricus Reference MüllerMüller, p. 450, pl. 14, figs. 1, 2, text-fig. 4.
1971Oneotodus cambricus (Müller); Reference MüllerMüller, pl. 2, fig. 3.
1979Muellerina cambricus (Müller); Reference BednarczykBednarczyk, p. 429, pl. 2, fig. 8.
1981Muellerodus cambricus (Müller); Miller in Reference RobisonRobison, p. W112, fig. 64, 3a, b.
1982Muellerodus pomeranensis Reference AnAn, p. 138, pl. 9, figs. 6, 8 (not 7).
1991Muellerodus cambricus (Müller); Reference Müller and HinzMüller and Hinz, p. 29, pl. 18, figs. 1–4, 6–13, 15, 17, text-fig. 10 (A, B).
2006Muellerodus cambricus (Müller); Reference Qi, Bagnoli and WangQi et al., p. 186, pl. 2, fig. 14.
2009Muellerodus cambricus (Müller); Reference Lee, Lee, Miller and JeongLee et al., p. 427, fig. 9 (3).
2011Muellerodus cambricus (Müller); Reference Bagnoli and QiBagnoli and Qi, p. 12, fig. 4 (J).
Materials.—
Six specimens.
Occurrence.—
Known from the Wangcun and Wa’ergang sections, where it ranges from the Westergaardodina lui-Westergaardodina ani Zone through the Eoconodontus Zone.
Remarks.—
The present specimens agree well with Muellerodus cambricus described and figured by Müller and Hinz (Reference Müller and Hinz1991).
Muellerodus guttulus Müller and Hinz, Reference Müller and Hinz1991
1991Muellerodus guttulus Reference Müller and HinzMüller and Hinz, p. 29, pl. 19, figs. 1–9, 11, text-fig. 10 (D).
Materials.—
Two specimens.
Occurrence.—
Known from the Wangcun section, where it occurs in the Westergaardodina lui-Westergaardodina ani Zone.
Remarks.—
The specimens at hand agree well with those described and figured by Müller and Hinz (Reference Müller and Hinz1991).
Muellerodus? obliquus (An, Reference An1982)
1982Proscandodus obliquus Reference AnAn, p. 148, pl. 9, figs. 1–5.
1993Muellerodus? obliquus (An); Reference DongDong, pl. 5, figs. 3, 11, 17.
2001aMuellerodus? obliquus (An); Dong and Bergström, p. 968, pl. 4, fig. 10.
2006Muellerodus? obliquus (An); Reference Qi, Bagnoli and WangQi et al., p. 186, pl. 2, figs. 10–11.
Materials.—
48 specimens.
Occurrence.—
Known from the Paibi and Wangcun sections, where it ranges from the Shandongodus priscus-Hunanognathus tricuspidatus Zone through the Proconodontus Zone.
Remarks.—
The present specimens are closely similar to An’s species. Müller and Hinz (Reference Müller and Hinz1991) considered Proscandodus to be a junior synonym of Furnishina. Our specimens are characterized by convex anterior and posterior sides, slender cusps, and one lateral costa that is more prominent than the other. These are features more characteristic of Muellerodus than Furnishina, and hence we refer this species to the former genus.
Muellerodus? oelandicus (Müller, Reference Müller1959)
1959Scandodus oelandicus Reference MüllerMüller, p. 463, pl. 12, fig. 14, 15, text-fig. 10.
1983Muellerodus oelandicus (Müller); Reference An, Zhang, Xiang, Zhang, Xu, Zhang, Jiang, Yang, Lin, Cui and YangAn et al., p. 109, pl. 3, fig. 9.
1986Proscandodus oelandicus (Müller); Reference Chen and GongChen and Gong, p. 171, pl. 34, fig. 13, text-fig. 67 (2).
1991Muellerodus? oelandicus (Müller); Reference Müller and HinzMüller and Hinz, p. 29, pl. 20, figs. 1–13, fig. 10 (E, F).
1993Muellerodus? oelandicus (Müller); Reference DongDong, pl. 3, figs. 6, 11.
2001aMuellerodus? oelandicus (Müller); Reference Dong and BergströmDong and Bergström, p. 968, pl. 5, fig. 12, pl. 6, fig. 9.
2006Muellerodus? oelandicus (Müller); Reference Qi, Bagnoli and WangQi et al., p. 186, pl. 2, fig. 12.
2009Muellerodus? oelandicus (Müller); Reference Lee, Lee, Miller and JeongLee et al., p. 423, fig. 7 (10).
2011Muellerodus? oelandicus (Müller); Reference Bagnoli and QiBagnoli and Qi, p. 12, fig. 4 (K).
Materials.—
32 specimens.
Occurrence.—
Known from Paibi, Wa’ergang and Wangcun sections, where it ranges from the Gapparodus bisulcatus-Westergaardodina brevidens Zone through the Cordylodus intermedius Zone.
Remarks.—
The specimens at hand agree closely with Müller’s species.
Muellerodus pomeranensis (Szaniawski, Reference Szaniawski1971)
1971Muellerina pomeranensis Reference SzaniawskiSzaniawski, p. 408, pl. 1, fig. 2, pl. 2, fig. 3, pl. 4, figs. 1–4.
1981Muellerodus pomeranensis (Szaniawski); Reference Miller, Sundberg, Harma and WrightMiller et al., fig. 4 (I, J).
1982Muellerodus pomeranensis (Szaniawski); Reference AnAn, p. 138, pl. 9, fig. 7 only (not figs. 6, 8), pl. 16, fig. 7, pl. 17, figs. 1, 3. (part)
1983Muellerodus pomeranensis (Szaniawski); Reference An, Zhang, Xiang, Zhang, Xu, Zhang, Jiang, Yang, Lin, Cui and YangAn et al., p. 110, pl. 3, fig. 10.
1987Muellerodus pomeranensis (Szaniawski); Reference AnAn, p. 108, pl. 3, figs. 4, 17.
1991Muellerodus pomeranensis (Szaniawski); Reference Müller and HinzMüller and Hinz, p. 30, pl. 18, figs. 5, 14, 16, 18–21, text-fig. 10 (C).
1993Muellerodus pomeranensis (Szaniawski); Reference DongDong, pl. 2, fig. 8.
2001aMuellerodus pomeranensis (Szaniawski); Reference Dong and BergströmDong and Bergström, p. 968, pl. 3, fig. 12.
2006Muellerodus pomeranensis (Szaniawski); Reference Qi, Bagnoli and WangQi et al., p. 186, pl. 2, fig. 13.
2009Muellerodus pomeranensis (Szaniawski); Reference Lee, Lee, Miller and JeongLee et al., p. 425, fig. 8 (3).
Materials.—
24 specimens.
Occurrence.—
Known from the Paibi and Wangcun sections where it ranges from the upper part of the Shandongodus priscus-Hunanognathus tricuspidatus Zone through the Westergaardodina cf. calix-Prooneotodus rotundatus Zone.
Remarks.—
The specimens at hand agree closely with previously described elements of this species, except that the flank costae of some specimens are indistinct. We do not feel that this slight difference merits taxonomic separation of our specimens from Muellerodus pomeranensis.
Genus Nogamiconus Miller, Reference Miller1980
Type species.—
Proacodus? sinensis (Nogami, Reference Nogami1966).
Nogamiconus sinensis (Nogami, Reference Nogami1966)
1966Proacodus? sinensis Reference NogamiNogami, p. 356, pl. 10, figs. 12–14.
1971Proacodus sinensis Nogami; Reference MüllerMüller, fig. 1c.
1981Nogamiconus sinensis (Nogami); Miller in Reference RobisonRobison, p. W112, fig. 64 (6a, b).
1982Gen et sp. indet. Reference AnAn, p. 156, pl. 8, fig. 7.
1983Nogamiconus sinensis (Nogami); Reference An, Zhang, Xiang, Zhang, Xu, Zhang, Jiang, Yang, Lin, Cui and YangAn et al., p. 110, pl. 30, fig. 11.
1991Nogamiconus sinensis (Nogami); Reference Müller and HinzMüller and Hinz, p. 31, pl. 21, figs. 1–6, text-fig. 12 (A, B).
1993Nogamiconus sinensis (Nogami); Reference DongDong, pl. 4, figs. 7–9.
2001aNogamiconus sinensis (Nogami); Reference Dong and BergströmDong and Bergström, p. 969, pl. 4, figs. 11–12.
Materials.—
178 specimens.
Occurrence.—
Known from Paibi and Wangcun sections, where it ranges from the Shandongodus priscus-Hunanognathus tricuspidatus Zone through the Westergaardodina lui-Westergaardodina ani Zone.
Remarks.—
The present specimens compare well with previous descriptions and illustrations of this species. Our collections includes several clusters containing only one type of element.
Genus Proacodus Müller, Reference Müller1959
Type species.—
Proacodus obliquus Müller, Reference Müller1959.
Proacodus obliquus Müller, Reference Müller1959
Figure 31.11–31.15, 31.17, 31.20–31.22, 31.25

Figure 31 (1–4) Paibiconus proarcuatus Dong, Reference Dong1993; (1) Chefu Formation, Wangcun section, GMPKU2769, posterolateral view; (2) Chefu Formation, Wangcun section, GMPKU2770, anterior view; (3) Huaqiao Formation, Wangcun section, GMPKU2002, anterior view; (4) Close-up view of the broken surface of (3), showing protoconodont structure; (5–10) Phakelodus tenuis (Müller, Reference Müller1959); (5) Chefu Formation, Wangcun section, GMPKU2771, cluster; (6) Bitiao Formation, Wangcun section, GMPKU2772, cluster; (7) Bitiao Formation, Wangcun section, GMPKU2773, lateral view; (8) Bitiao Formation, Wangcun section, GMPKU2774, cluster; (9) Shenjiawan Formation, Wa’ergang section, GMPKU2775, lateral view; (10) Shenjiawan Formation, Wa’ergang section, GMPKU2776, cluster; (11–15, 17, 20–22, 25) Proacodus obliquus Müller, Reference Müller1959; (11) Bitiao Formation, Wangcun section, GMPKU2777, top view; (12) Bitiao Formation, Wangcun section, GMPKU2778, posterior view; (13) Bitiao Formation, Wangcun section, GMPKU2779, posterior view; (14) Bitiao Formation, Wangcun section, GMPKU2780, posterior view; (15) Bitiao Formation, Wangcun section, GMPKU2781, posterior view; (17) Bitiao Formation, Wangcun section, GMPKU2782, posterior view; (20) Bitiao Formation, Wangcun section, GMPKU2783, posterior view; (21) Bitiao Formation, Wangcun section, GMPKU2784, top view from anterior side; (22) Bitiao Formation, Wangcun section, GMPKU2785, posterior view; (25) Shenjiawan Formation, Wa’ergang section, GMPKU2786, top view from anterior side; (16, 18, 19, 23, 24, 26) Proacodus pulcherus (An, Reference An1982); (16) Bitiao Formation, Wangcun section, GMPKU2787, posterior view; (18) Bitiao Formation, Wangcun section, GMPKU2788, posterior view; (19) Bitiao Formation, Wangcun section, GMPKU2789, posterior view; (23) Bitiao Formation, Wangcun section, GMPKU2790, posterior view; (24) Bitiao Formation, Wangcun section, GMPKU2791, posterior view; (26) Bitiao Formation, Wangcun section, GMPKU2792, posterior view. Relative scale bar represents 311 μm (1), 362 μm (2), 227 μm (3), 71 μm (4), 130 μm (5, 8, 9, 12–26), 188 μm (6), 160 μm (7), 119 μm (10), 100 μm (11).
1959Proacodus obliquus Reference MüllerMüller, p. 458, pl. 13, figs. 1, 4 (not fig. 2). (part)
1971Proacodus obliquus Müller; Reference MüllerMüller, pl. 2, fig. 2.
1979Proacodus obliquus Müller; Reference BednarczykBednarczyk, p. 433, pl. 1, figs. 11, 14.
1981Proacodus obliquus Müller; Miller in Reference RobisonRobison, p. W112, fig. 64 (4a, b).
1991Proacodus obliquus Müller; Reference Müller and HinzMüller and Hinz, p. 34, pl. 22, figs. 12–23, fig. 13 (C).
Description.—
Proclined, asymmetrical elements with a long lateral process. Both anterior and posterior sides are broadly rounded. The lateral process is very long and keeled on its upper side. Basal cavity is large and deep, the apex of which extends past the point of flexure of the element. Cross section is round at the apex and becomes an elongated oval at the base.
Materials.—
30 specimens.
Occurrence.—
Known from the Wangcun and Wa’ergang sections, where it ranges from the Westergaardodina lui-Westergaardodina ani Zone through the Eoconodontus Zone.
Proacodus pulcherus (An, Reference An1982)
Figures 23.1–23.6, 31.16, 31.18, 31.19, 31.23, 31.24, 31.26
1959Proacodus obliquus Reference MüllerMüller, p. 458, pl. 13, fig. 2 only. (part)
1982Muellerodus pulcherus Reference AnAn, p. 139, pl. 9, figs. 13, 15, pl. 10, figs. 12, 14.
1991Proacodus pulcherus (An); Reference Müller and HinzMüller and Hinz, p. 34, pl. 22, figs. 3–5, 7–11, text-fig. 13 (A, B).
2000Proacodus obliquus Müller; Reference Pyle and BarnesDubinina, p. 172, pl. 3, figs. 4–6.
2006Proacodus pulcherus (An); Reference Qi, Bagnoli and WangQi et al., p. 187, pl. 3, fig. 18.
2011Proacodus pulcherus (An); Reference Bagnoli and QiBagnoli and Qi, p. 12, fig. 4 (L).
Description.—
Coniform elements, proclined, asymmetrical, with a short lateral process. Both anterior and posterior sides are broadly rounded. The lateral process is short and keeled on its upper side. Basal cavity is large and deep, with the apex extending past the point of flexure of the element. Cross section is round at the apex and becomes tear-shaped at the base.
Materials.—
40 specimens.
Occurrence.—
Known from the Wangcun and Wa’ergang sections, where it ranges from the Westergaardodina lui-Westergaardodina ani Zone through the Proconodontus Zone.
Proacodus cf. pulcherus (An, Reference An1982)
cf.1959 Proacodus obliquus Reference MüllerMüller, p. 458, pl. 13, fig. 2, only. (part)
cf.1982 Muellerodus pulcherus Reference AnAn, p. 139, pl. 9, figs. 13, 15, pl. 10, figs. 12, 14.
cf.1991 Proacodus pulcherus (An); Reference Müller and HinzMüller and Hinz, p. 34, pl. 22, figs. 3–5, 7–11, text-fig. 13 (A, B).
Description.—
Coniform elements, proclined, asymmetrical, with a long lateral process. Both anterior and posterior sides are broadly rounded. The long lateral process extends horizontally and then upward and has a keel on its upper side. Basal cavity large and deep, with the apex extending past the point of flexure of the element. Cross section is round at the apex and becomes tear-shaped at the base.
Materials.—
Three specimens.
Occurrence.—
Known from Wa’ergang sections, where it occurs in the Proconodontus Zone.
Remarks.—
This form is similar to Proacodus pulcherus, but it differs in its longer process, which extends upward. Also, the apex of the present species is straight, whereas the apex of P. pulcherus is twisted.
Genus Prooneotodus Müller and Nogami, Reference Müller and Nogami1971
Type species.—
Oneotodus gallatini Müller, Reference Müller1959.
Prooneotodus gallatini (Müller, Reference Müller1959)
Figures 24.9, 24.10, 24.12–24.32, 32.1–32.15, 32.17–32.21

Figure 32 (1–15, 17–21) Prooneotodus gallatini Müller, Reference Müller1959; (1) Shenjiawan Formation, Wa’ergang section, GMPKU2856, lateral view; (2) Shenjiawan Formation, Wa’ergang section, GMPKU2857, lateral view; (3) Shenjiawan Formation, Wa’ergang section, GMPKU2858, lateral view; (4) Shenjiawan Formation, Wa’ergang section, GMPKU2859, lateral view; (5) Shenjiawan Formation, Wa’ergang section, GMPKU2860, lateral view; (6) Shenjiawan Formation, Wa’ergang section, GMPKU2861, lateral view; (7) Shenjiawan Formation, Wa’ergang section, GMPKU2862, oblique posterior view from base; (8) Shenjiawan Formation, Wa’ergang section, GMPKU2863, lateral view; (9) Shenjiawan Formation, Wa’ergang section, GMPKU2864, lateral view; (10) Shenjiawan Formation, Wa’ergang section, GMPKU2865, lateral view; (11) Shenjiawan Formation, Wa’ergang section, GMPKU2866, lateral view; (12) Shenjiawan Formation, Wa’ergang section, GMPKU2867, lateral view; (13) Shenjiawan Formation, Wa’ergang section, GMPKU2868, lateral view; (14) Shenjiawan Formation, Wa’ergang section, GMPKU2869, lateral view; (15) Shenjiawan Formation, Wa’ergang section, GMPKU2870, lateral view; (17) Shenjiawan Formation, Wa’ergang section, GMPKU2871, lateral view; (18) Shenjiawan Formation, Wa’ergang section, GMPKU2872, lateral view; (19) Shenjiawan Formation, Wa’ergang section, GMPKU2873, lateral view; (20) Shenjiawan Formation, Wa’ergang section, GMPKU2874, lateral view; (21) Shenjiawan Formation, Wa’ergang section, GMPKU2875, lateral view; (16, 22–29) Prooneotodus rotundatus (Druce and Jones, Reference Druce and Jones1971); (16) Bitiao Formation, Wangcun section, GMPKU2876, lateral view; (22) Shenjiawan Formation, Wa’ergang section, GMPKU2877, lateral view; (23) Shenjiawan Formation, Wa’ergang section, GMPKU2878, lateral view; (24) Shenjiawan Formation, Wa’ergang section, GMPKU2879, cluster, lateral view; (25) Shenjiawan Formation, Wa’ergang section, GMPKU2880, lateral view; (26) Shenjiawan Formation, Wa’ergang section, GMPKU2881, lateral view; (27) Shenjiawan Formation, Wa’ergang section, GMPKU2882, lateral view; (28) Shenjiawan Formation, Wa’ergang section, GMPKU2883, lateral view; (29) Shenjiawan Formation, Wa’ergang section, GMPKU2884, lateral view. Relative scale bar represents 130 μm (1–13, 16–29), 138 μm (14, 15).
1959Oneotodus gallatini Reference MüllerMüller, p. 457, pl. 13, figs. 5–10, 12, 18.
1971Oneotodus gallatini Müller; Reference Druce and JonesDruce and Jones, p. 81, pl. 9, figs. 9a–c, pl. 10, figs. 9a–10c, text-figs. 26f, g.
1973Oneotodus gallatini Müller; Reference Özgül and GedikÖzgül and Gedik, p. 48, pl. 1, figs. 1, 20.
1980Oneotodus gallatini Müller; Lee, pl. 1, fig. 4.
1981Prooneotodus gallatini (Müller); Miller in Reference RobisonRobison, p. W113, fig. 64 (2a–c).
1981Prooneotodus gallatini (Müller); Reference Miller, Sundberg, Harma and WrightMiller et al., fig. 4K–M.
1982Prooneotodus gallatini (Müller); Reference AnAn, p. 144, pl. 11, figs. 5–6, 9–14, pl. 16, fig. 13.
1982Prooneotodus gallatini (Müller); Reference Fortey, Landing and SkevingtonFortey et al., p. 125, text-figs. 9 (H, O).
1983Prooneotodus gallatini (Müller); Reference An, Zhang, Xiang, Zhang, Xu, Zhang, Jiang, Yang, Lin, Cui and YangAn et al., p. 128, pl. 5, fig. 7.
1983Prooneotodus gallatini (Müller); Reference AzmiAzmi, p. 379, pl. 3, figs. 3, 5.
1985Prooneotodus gallatini (Müller); Reference WangWang, p. 235, pl. 3, figs. 23–25, pl. 5, figs. 4, 5, pl. 8, figs. 12, 21, pl. 12, figs. 22, 26, pl. 13, fig. 10, pl. 14, fig. 5.
1986“Prooneotodus” gallatini (Müller); Reference Chen and GongChen and Gong, p. 166, pl. 22, figs. 13, 17 (not 15, 16), pl. 23, figs. 2–3, 7, 10, 16–19, pl. 24, fig. 12, text-figs. 64 (2, 5, not 3). (part)
1987Prooneotodus gallatini (Müller); Reference AnAn, p. 112, pl. 2, figs. 1, 2, 5, 6, 11, 15.
1987Prooneotodus gallatini (Müller); Reference Buggisch and RepetskiBuggisch and Repetski, p. 159, pl. 8, fig. 2.
1991Prooneotodus gallatini (Müller); Reference Müller and HinzMüller and Hinz, p. 37, pl. 24, figs. 1–28, 29.
2000Prooneotodus gallatini (Müller); Reference Pyle and BarnesDubinina, p. 178, pl. 4, figs. 22, 29, 30.
2000“Prooneotodus” mitriformis Reference Pyle and BarnesDubinina, p. 178, pl. 4, fig. 28.
2000Prooneotodus rotundatus (Druce et Jones); Reference Zhao, Zhang and XiaoZhao et al., p. 216, pl. 23, figs. 25, 28.
2002aProoneotodus gallatini (Müller); Reference LeeLee, p. 165, pl. 2, fig. 10.
2002bProoneotodus gallatini (Müller); Reference LeeLee, p. 27, pl. 2, fig. 7.
2004Prooneotodus gallatini (Müller); Reference LeeLee, p. 60, fig. 5 (N).
2007Prooneotodus gallatini (Müller); Reference Landing, Westrop and KeppieLanding et al., p. 922, fig. 9 (d, e).
2009Prooneotodus gallatini (Müller); Reference Lee, Lee, Miller and JeongLee et al., p. 423, fig. 7 (12, 15), p. 425, fig. 8 (16), p. 427, fig. 9 (2).
2011Prooneotodus gallatini (Müller); Reference Bagnoli and QiBagnoli and Qi, p. 12, fig. 4 (C).
Description.—
Moderate to small coniform elements, erect to proclined, symmetrical to asymmetrical. The apex is pointed. Both anterior and posterior sides are rounded. No keels or costae are present. Basal cavity is deep, with the apex extending past the point of flexure of the element or even nearly to its tip. The cross section is round at the apex and changes to variably oval at the base.
Materials.—
300 specimens.
Occurrence.—
Known from the Wangcun and Wa’ergang sections, where it ranges from the Westergaardodina lui-Westergaardodina ani Zone through the Cordylodus intermedius Zone.
Prooneotodus rotundatus (Druce and Jones, Reference Druce and Jones1971)
1971Coelocerodontus rotundatus Reference Druce and JonesDruce and Jones, p. 62, pl. 9, figs. 10–13, text-figs. 22c, d.
1973Proconodontus cf. rotundatus (Druce and Jones); Reference MüllerMüller, p. 43, pl. 3, figs. l–3.
1976Prooneotodus rotundatus (Druce and Jones); Reference Miller and PadenMiller and Paden, p. 596, pl. 1, figs. 24, 25.
1982Prooneotodus rotundatus (Druce and Jones); Reference AnAn, p. 144, pl. 4, fig. 12, pl. 11, figs. l–4, 7, 8.
1982Prooneotodus rotundatus (Druce and Jones); Reference Fortey, Landing and SkevingtonFortey et al., p. 125, text-fig. 9 (J, M).
1983Prooneotodus rotundatus (Druce and Jones); Reference An, Zhang, Xiang, Zhang, Xu, Zhang, Jiang, Yang, Lin, Cui and YangAn et al., p. 129, pl. 5, fig. 6.
1985Prooneotodus rotundatus (Druce and Jones); Reference WangWang, p. 236, pl. 3, figs. 5–7, pl. 6, fig, 13, pl. 7, figs. 7, 8, pl. 8, fig. 6, pl. 9, fig. 2, pl. 10, fig. 9, pl. 12, fig. 6, pl. 13, figs. 8, ?32.
1986“Prooneotodus” rotundatus (Druce and Jones); Reference Chen and GongChen and Gong, p. 167, pl. 22, figs. 1–4 (not 5), 6, 9–11, 18, 20, pl. 23, figs. 1, 9, 12, 14, 20 (not pl. 26, fig. 9), text-fig. 64 (1). (part)
1987Prooneotodus rotundatus (Druce and Jones); s.f. Reference DongDong, p. 170, pl. 1, fig. 29, text-fig. 3M.
2000Prooneotodus rotundatus (Druce and Jones); Reference Pyle and BarnesDubinina, p. 179, pl. 4, figs. 26, 27.
2001Prooneotodus rotundatus (Druce and Jones); Reference LeeLee, p. 449, fig. 6 (5).
2002Prooneotodus rotundatus (Druce and Jones); Lee, p. 165, pl. 2, fig. 8.
2002Prooneotodus rotundatus (Druce and Jones); Lee, p. 27, pl. 2, fig. 8.
2004Prooneotodus rotundatus (Druce and Jones); Reference LeeLee, p. 60, fig. 5 (I).
2009Prooneotodus rotundatus (Druce and Jones); Reference Lee, Lee, Miller and JeongLee et al., p. 427, fig. 9 (1).
Description.—
Coniform elements, moderate to small, proclined and symmetrical. The apex is pointed. Both anterior and posterior sides are rounded. No keels or costae are present. Basal cavity is very deep, with the apex extending nearly to the tip of the element. The cross section is circular through the small cusp and base.
Materials.—
90 specimens.
Occurrence.—
Known from the Wangcun and Wa’ergang sections, where it ranges from Westergaardodina cf. calix-Prooneotodus rotundatus Zone through the Cordylodus intermedius Zone.
Remarks.—
Prooneotodus rotundatus differs from Prooneotodus gallatini in its circular cross section and deeper basal cavity. In addition, elements of the former taper gradually, whereas elements of the latter taper relatively rapidly. In gross morphology, P. rotundatus resembles Millerodontus intermedius n. gen. n. sp., but the latter is recognized as a primitive euconodont in the light of histological investigation. Accordingly, some published specimens previously assigned to P. rotundatus probably should be reassigned to M. intermedius, but this could only be done by means of histological investigation.
Prooneotodus terashimai (Nogami, Reference Nogami1967)

Figure 33 (1–8) Prooneotodus terashimai (Nogami, Reference Nogami1967); (1) Bitiao Formation, Wangcun section, GMPKU2885, lateral view; (2) Bitiao Formation, Wangcun section, GMPKU2886, lateral view; (3) Bitiao Formation, Wangcun section, GMPKU2887, lateral view; (4) Bitiao Formation, Wangcun section, GMPKU2888, lateral view; (5) Bitiao Formation, Wangcun section, GMPKU2889, lateral view; (6) Bitiao Formation, Wangcun section, GMPKU2890, lateral view; (7) Bitiao Formation, Wa’ergang section, GMPKU2891, lateral view; (8) Shenjiawan Formation, Wa’ergang section, GMPKU2892, lateral view; (9) Prooneotodus? sp. A, Shenjiawan Formation, Wa’ergang section, GMPKU2893, posterior view; (10, 11) Prosagittodontus compressus n. sp.; (10) Chefu Formation, Wangcun section, holotype, GMPKU2894, anterior view; (11) Chefu Formation, Wangcun section, GMPKU2895, posterior view; (12–28) Prosagittodontus dalmani (Müller, Reference Müller1959); (12) Bitiao Formation, Wangcun section, GMPKU2896, posterior view; (13) Bitiao Formation, Wangcun section, GMPKU2897, posterior view; (14) Bitiao Formation, Wangcun section, GMPKU2898, posterior view; (15) Bitiao Formation, Wangcun section, GMPKU2899, posterior view; (16) Bitiao Formation, Wangcun section, GMPKU2900, posterior view; (17) Bitiao Formation, section, Wa’ergang GMPKU2901, posterior view; (18) Shenjiawan Formation, Wa’ergang section, GMPKU2902, oblique posterior view; (19) Bitiao Formation, Wangcun section, GMPKU2903, posterior view; (20) Bitiao Formation, Wangcun section, GMPKU2904, posterior view; (21) Bitiao Formation, Wangcun section, GMPKU2905, posterior view; (22) Bitiao Formation, Wangcun section, GMPKU2906, oblique posterior view; (23) Shenjiawan Formation, Wa’ergang section, GMPKU2907, posterior view; (24) Shenjiawan Formation, Wa’ergang section, GMPKU2908, posterior view; (25) Shenjiawan Formation, Wa’ergang section, GMPKU2909, posterior view; (26) Shenjiawan Formation, Wa’ergang section, GMPKU2910, posterior view; (27) Shenjiawan Formation, Wa’ergang section, GMPKU2911, posterior view; (28) Shenjiawan Formation, Wa’ergang section, GMPKU2912, oblique posterior view. Relative scale bar represents 137 μm (1–19, 21–28), 144 μm (20).
1967Oneotodus terashimai Reference NogamiNogami, p. 217, pl. 1, figs. 14–16.
1985Prooneotodus terashimai cylindricum Reference An and HuAn and Hu, pl. 1, fig. 1.
1987Prooneotodus terashimai cylindricum An and Hu; Reference AnAN, p. 113, pl. 1, figs. 1–3.
2000Prooneotodus terashimai kazakhstanica Reference Pyle and BarnesDubinina, p. 179, pl. 4, figs. 19, 19a, 20, 20a.
2000Phakelodus terashimai (Nogami); Reference Göncüoğlu and KozurGöncüoğlu and Kozur, p. 599, fig. 4 (d).
? 2009Prooneotodus terashimai (Nogami); Reference Lee, Lee, Miller and JeongLee et al., p. 427, fig. 9 (11).
Description.—
Coniform elements, moderate to small, and symmetrical. The cusp is small and hook-like. The base is slender and cylindrical. Both anterior and posterior edges are rounded. No keels or costae are present. Basal cavity is very deep, and the apex extends past the point of flexure of the element.
Materials.—
20 specimens.
Occurrence.—
Known from the Wangcun and Wa’ergang sections, where it ranges from the Westergaardodina lui-Westergaardodina ani Zone through the Cordylodus proavus Zone.
Remarks.—
Prooneotodus terashimai is characterized by the minute and hook-like cusp. An and Hu (Reference An and Hu1985) erected a new subspecies Prooneotodus terashimai cylindricum. The only difference between them is that the latter is much slender in gross morphology. It is considered as intraspecific variation herein.
Prooneotodus? sp. A
Description.—
Large coniform element with pointed apex. There is no conspicuous junction between cusp and base. The anterior side is slightly convex, whereas the posterior side is strongly convex. Keels and costae are lacking. The cross-section is circular at the apex and becomes roundly triangular in the rest of the element. Basal cavity is large and extends nearly to tip.
Materials.—
Only one specimen.
Occurrence.—
Known from the Wa’ergang section, where it occurs in the Eoconodontus Zone.
Genus Prosagittodontus Müller and Nogami, Reference Müller and Nogami1971
Type species.—
Sagittodontus dalmani Müller, Reference Müller1959.
Prosagittodontus compressus new species
1994Laiwugnathus sagittatus Reference An and MeiAn and Mei, p. 537, pl. 1, figs. 28, 29.
2001aProsagittodontus? sp. Dong and Bergström, p. 963, pl. 1, figs. 17, 18.
Diagnosis.—
A species of Prosagittodontus with two lateral projections inclining downward, with the outer parts strongly compressed. A third projection becomes a rounded median keel or an indistinct bulge. The basal cavity is moderately large and deep.
Description.—
Coniform elements shaped like arrowheads in outline. The apical angle is less than 90°. From the pointed tip, the flanks diverge in an almost straight line, separating into two lateral projections that are strongly compressed in the outer parts, with sharp costae on the margins. A third projection becomes a rounded median keel or an indistinct bulge. The anterior side of elements is flattened, whereas the posterior side is concave. The basal cavity is moderately large and deep.
Etymology.—
compressus, Latin, compressed.
Types.—
Holotype: GMPKU2894, from the Westergaardodina matsushitai-Westergaardodina grandidens Zone, Chefu Formation, Wangcun section, Wangcun village, Yongshun county, Hunan Province.
Materials.—
Nine specimens.
Occurrence.—
Known from the Wangcun and Paibi sections, where it ranges from the Westergaardodina quadrata Zone through the Westergaardodina lui-Westergaardodina ani Zone.
Remarks.—
Prosagittodontus compressus n. sp. appears to be the transitional form among Prosagittodontus, Laiwugnathus, and tricuspidate Westergaardodina. However, the outline of Laiwugnathus is triangular or subtriangular, while tricuspidate Westergaardodina has a median projection. Accordingly, this species is assigned to Prosagittodontus herein.
Prosagittodontus dalmani (Müller, Reference Müller1959)
1959Sagittodontus dalmani Reference MüllerMüller, p. 460, pl. 14, figs. 5, 7, 10.
1971Sagittodontus dalmani Müller; Reference MüllerMüller, pl. 1, fig. 8.
1978Prosagittodontus dalmani (Müller); Reference AbaimovaAbaimova, p. 501, pl. 8, figs. 3, 5, 7.
1981Prosagittodontus dalmani (Müller); Miller in Reference RobisonRobison, p. W114, fig. 65. 2a, b.
1982Prosagittodontus dalmani (Müller); Reference AnAn, p. 147, pl. 5, fig. 6.
1986Prosagittodontus dalmani (Müller); Reference Chen, Zhang and YuChen et al., p. 369, pl. 2, figs. 14, 15.
1987Prosagittodontus dalmani (Müller); Reference AnAn, p. 113, pl. 3, figs. 23, 24.
1991Prosagittodontus dalmani (Müller); Reference Müller and HinzMüller and Hinz, p. 37, pl. 25, figs. 1–22, text-fig. 14 (B).
2002Prosagittodontus dalmani (Müller); Lee, p. 25, pl. 1, fig. 15.
2009Prosagittodontus dalmani (Müller); Reference Lee, Lee, Miller and JeongLee et al., p. 427, fig. 9.6.
Materials.—
90 specimens.
Occurrence.—
Known from the Wangcun and Paibi sections, where it ranges from the Westergaardodina matsushitai-Westergaardodina grandidens Zone through the Cordylodus proavus Zone.
Remarks.—
The specimens at hand agree well with those described and figured by Müller (Reference Müller1959) and Müller and Hinz (Reference Müller and Hinz1991). The median projection of Prosagittodontus dalmani is prominent and extends posteriorly, resulting in a crudely triangular basal cross-section of the element. So, P. dalmani differs from Prosagittodontus compressus.
Prosagittodontus cf. eureka (Müller, Reference Müller1959)
1959 cf.Sagittodontus eureka Reference MüllerMüller, p. 461, pl. 14, fig. 6.
1973 cf.Prosagittodontus eureka (Müller); Reference MüllerMüller, p. 45, pl. 5, figs. 1–2.
1982Prosagittodontus eureka (Müller); Reference AnAn, p. 147, pl. 5, figs. 1–2.
1982Prosagittodontus aff. eureka (Müller); Reference AnAn, p. 148, pl. 5, figs. 4, 8.
2011Prosagittodontus aff. eureka (Müller); Reference Bagnoli and QiBagnoli and Qi, p. 12, fig. 4 (D, E).
Materials.—
Four specimens.
Occurrence.—
Known from Wangcun section, where it occurs Westergaardodina lui-Westergaardodina ani Zone.
Remarks.—
The specimens at hand are similar to those described and figured by Müller (Reference Müller1959, Reference Müller1973), but the median keel on the posterior side of the present specimens is much less distinct than that of Prosagittodontus eureka s. s. An (Reference An1982) described and figured four specimens assigned to P. eureka or Prosagittodontus aff. eureka. Those specimens should be all assigned to Prosagittodontus cf. eureka.
Genus Serratocambria Müller and Hinz, Reference Müller and Hinz1991
Type species.—
Serratocambria minuta Müller and Hinz, Reference Müller and Hinz1991.
Serratocambria minuta Müller and Hinz, Reference Müller and Hinz1991
1991Serratocambria minuta Reference Müller and HinzMüller and Hinz, p. 38, pl. 27, figs. 1–17, text-fig. 15 (A, B).
1994Serratocambria dacryoidea Reference An and MeiAn and Mei, p. 538, pl. 2, figs. 25–26.
Description.—
Ramiform elements, small and asymmetrical, with a long serrated lateral process. The small cusp is strongly recurved and spine-like. Both anterior and posterior sides are broadly rounded. The lateral process is long, compressed, and serrated on its upper edge. Basal cavity is moderate in size, with the apex extending nearly up to the point of flexure of the element. Cross section is round at the apex and changes into a variable outline at the base.
Materials.—
12 specimens.
Occurrence.—
Known only from the Wangcun section, where it occurs in the Westergaardodina cf. calix-Prooneotodus rotundatus Zone.
Remarks.—
An and Mei (Reference An and Mei1994) erected a new species, Serratocambria dacryoidea. It differs from Serratocambria minuta in its short lateral process and the erect, serrated tiny denticles. The short lateral process is broken, and the shape of the denticles is considered to represent intraspecific variation. Thus, the two species are considered synonyms herein.
Genus Shandongodus An, Reference An1982
Type species.—
Shandongodus priscus An, Reference An1982.
Shandongodus priscus An, Reference An1982
1982Shandongodus priscus Reference AnAn, p. 149, pl. 4, figs. 5–8, pl. 5, fig. 10.
1990Shandongodus priscus An; Dong, pl. 1, figs. 10, 12, 13.
1993Shandongodus priscus An; Reference DongDong, pl. 3, fig. 4.
1994Shandongodus priscus An; Reference An and MeiAn and Mei, pl. 2, figs. 17, 24.
2001aShandongodus priscus An; Reference Dong and BergströmDong and Bergström, p. 972, pl. 3, fig. 14.
2008Shandongodus priscus An; Reference Bagnoli, Qi and WangBagnoli et al., p. 112, fig. 3 (F, G).
Materials.—
Eight specimens.
Occurrence.—
Known from Paibi and Wangcun sections, where it occurs in the Shandongodus priscus-Hunanognathus tricuspidatus Zone.
Remarks.—
The specimens at hand agree in all respects with those described and illustrated by An (Reference An1982).
Genus Wangcunognathus new genus
Type species.—
Wangcunognathus elegans n. gen. n. sp.
Diagnosis.—
Reclined coniform elements, with a short, adenticulate, posterior process which possesses a sharp oral costa with a conspicuous indentation. Anterior side of elements possess two sharp antero-lateral costae extending from apex to base. Posterior side of cusp possesses a sharp costa. Basal cavity deep, with a tip that ends higher than the top of posterior process.
Etymology.—
Named for the Wangcun section in Hunan Province and gnathus, Greek, jaw.
Wangcunognathus elegans new genus new species

Figure 34 (1–26) Wangcunognathus elegans n. gen. n. sp.; (1) Bitiao Formation, Wangcun section, GMPKU2941, lateral view; (2) Bitiao Formation, Wangcun section, GMPKU2942, lateral view; (3) Bitiao Formation, Wangcun section, GMPKU2943, lateral view; (4) Bitiao Formation, Wangcun section, GMPKU2944, lateral view; (5) Bitiao Formation, Wangcun section, GMPKU2945, lateral view; (6) Bitiao Formation, Wangcun section, GMPKU2946, lateral view; (7) Bitiao Formation, Wangcun section, GMPKU2947, lateral view; (8) Bitiao Formation, Wangcun section, GMPKU2948, lateral view; (9) Bitiao Formation, Wangcun section, GMPKU2949, lateral view; (10) Bitiao Formation, Wangcun section, holotype, GMPKU2950, lateral view; (11) Bitiao Formation, Wangcun section, GMPKU2951, lateral view; (12) Bitiao Formation, Wangcun section, GMPKU2952, lateral view; (13) Bitiao Formation, Wangcun section, GMPKU2953, lateral view; (14) Bitiao Formation, Wangcun section, GMPKU2954, posterior view; (15) Bitiao Formation, Wangcun section, GMPKU2955, lateral view; (16) Bitiao Formation, Wangcun section, GMPKU2956, lateral view; (17) Bitiao Formation, Wangcun section, GMPKU2957, oblique anterior view; (18) Bitiao Formation, Wangcun section, GMPKU2958, lateral view; (19) Bitiao Formation, Wangcun section, GMPKU2959, posterior view; (20) Bitiao Formation, Wangcun section, GMPKU2960, oblique anterior view; (21) Bitiao Formation, Wangcun section, GMPKU2961, lateral-anterior view; (22) Bitiao Formation, Wangcun section, GMPKU2962, lateral view; (23) Bitiao Formation, Wangcun section, GMPKU2963, lateral view; (24) Bitiao Formation, Wangcun section, GMPKU2964, lateral view; (25) Bitiao Formation, Wangcun section, GMPKU2965, lateral view; (26) Bitiao Formation, Wangcun section, GMPKU2966, lateral view. Scale bar represents 127 μm (1–26).
Diagnosis.—
As for the genus.
Description.—
Reclined coniform elements with a short adenticulate, posterior process. It is asymmetrical due to lateral bending. The anterior sides of elements are slightly convex with two sharp anterolateral costae extending from apex to base. The posterior sides of cusps possess a sharp costa, which leads to the subtriangular cross-section of the cusp. The base is slightly either compressed or expanded laterally to a variable extent, so the cross-section of the base is variable in shape. The oral margin is characterized by a sharp costa with a conspicuous indentation. Basal cavity deep, with a tip that ends above the top of the posterior process.
Etymology.—
elegans, Latin, elegant.
Types.—
Holotype: GMPKU2950 from the Westergaardodina lui-Westergaardodina ani Zone, Bitiao Formation, Wangcun section, Wangcun village, Yongshun county, Hunan Province.
Materials.—
300 specimens.
Occurrence.—
Known from the Wangcun section, where it occurs in the Westergaardodina lui-Westergaardodina ani Zone.
Remarks.—
Wangcunognathus elegans n. gen. n. sp. probably is a paraconodont.
Genus Westergaardodina Müller, Reference Müller1959
Type species.—
Westergaardodina bicuspidata Müller, Reference Müller1959.
Remarks.—
The morphology of Westergaardodina is so different from that of other conodonts that some completely different morphological terms are used to describe these species. For example, there are two groups of species of Westergaardodina, those with two cusp-like features (bicuspidate) and those with three (tricuspidate). These cusp-like features are referred to as projections, two of which are lateral projections and the third (if present) is a median projection. The lateral projections may have a basal cavity or not. There may be a depression or groove around part or all of the outer perimeter of the lateral projections, referred to as a lateral cavity. In some species the projections are directed upward from the lower part of the element but then bend outward in the upper part; the turning point is where the bending begins. The morphology of elements described herein is based on the terms introduced or modified by Müller and Hinz (Reference Müller and Hinz1991, p. 14–15).
Westergaardodina ahlbergi Müller and Hinz, Reference Müller and Hinz1991

Figure 35 (1) Westergaardodina sp. C, Shenjiawan Formation, Wa’ergang section, GMPKU2967, posterior view; (2, 4–6) Westergaardodina ani Dong, Repetski and Bergström, 2004c; (2) Bitiao Formation, Wangcun section, GMPKU2968, posterior view; (4) Bitiao Formation, Wangcun section, GMPKU2969, posterior view; (5) Bitiao Formation, Wangcun section, GMPKU2970, posterior view; (6) Panjiazui Formation, Wa’ergang section, GMPKU2971, posterior view; (3) Westergaardodina ahlbergi Müller and Hinz, Reference Müller and Hinz1991, Bitiao Formation, Wangcun section, GMPKU2972, posterior view; (7, 8) Westergaardodina brevidens Dong, Reference Dong1993; (7) Huaqiao Formation, Paibi section, GMPKU2973, posterior view; (8) Huaqiao Formation, Paibi section, GMPKU2974, posterior view; (9) Westergaardodina cf. behrae Müller and Hinz, Reference Müller and Hinz1991, Bitiao Formation, Wangcun section, GMPKU2975, posterior view; (10–20) Westergaardodina cf. calix Müller and Hinz, Reference Müller and Hinz1991; (10) Bitiao Formation, Wangcun section, GMPKU2136, posterior view; (11) Bitiao Formation, Wangcun section, GMPKU2135, posterior view; (12) Bitiao Formation, Wangcun section, GMPKU2137, posterior view; (13) Bitiao Formation, Wangcun section, GMPKU2130, posterior view; (14) Bitiao Formation, Wangcun section, GMPKU2134, posterior view; (15) Bitiao Formation, Wangcun section, GMPKU2128, posterior view; (16) Bitiao Formation, Wangcun section, GMPKU2127, posterior view; (17) Bitiao Formation, Wangcun section, GMPKU2129, posterior view; (18) Shenjiawan Formation, Wa’ergang section, GMPKU2133, posterior view; (19) Bitiao Formation, Wangcun section, GMPKU2131, posterior view; (20) Bitiao Formation, Wangcun section, GMPKU2132, posterior view; (21) Westergaardodina sp. D, Huaqiao Formation, Wangcun section, GMPKU2976, posterior view; (22–26) Westergaardodina dimorpha n. sp.; (22) Bitiao Formation, Wangcun section, GMPKU2977, posterior view; (23) Bitiao Formation, Wangcun section, GMPKU2978, posterior view; (24) Bitiao Formation, Wangcun section, GMPKU2979, posterior view; (25) Bitiao Formation, Wangcun section, holotype, GMPKU2980, posterior view; (26) Bitiao Formation, Wangcun section, GMPKU2981, anterior view. Relative scale bar represents 171 μm (1, 3, 6, 25, 26), 165 μm (2, 4, 5, 8–13, 15, 18, 20, 21, 23, 24), 156 μm (7, 14, 16, 17, 19), 264 μm (22).
? 1973Westergaardodina muelleri (Nogami); Reference MüllerMüller, p. 48, pl. 2, fig. 9.
1982Westergaardodina cf. muelleri (Nogami); Reference AnAn, p. 153, pl. 6, fig. 11.
1991Westergaardodina ahlbergi Reference Müller and HinzMüller and Hinz, p. 41, pl. 38, figs. 1–11.
Materials.—
Four specimens.
Occurrence.—
Known from the Wangcun section, where it occurs in the Westergaardodina lui-Westergaardodina ani Zone.
Remarks.—
The specimens at hand agree in all aspects with the species described and figured by Müller and Hinz (Reference Müller and Hinz1991).
Westergaardodina ani Dong, Repetski, and Bergström, 2004c
2004cWestergaardodina ani Dong, Repetski, and Bergström, p. 1197, pl. 3, figs. 11–13, 16, 21.
Description.—
Small, bicuspidate Westergaardodina. The anterior side is convex and characterized by deeply incised turning points, which may be indistinct due to the tiny, spine-like median projection. The median projection is absent in larger specimens. The posterior side is deeply concave. The two lateral projections are nearly of the same length, and the interspace between them is generally extremely narrow. The length/width ratio is nearly three to one. The gross morphology of the unit looks like a slender spoon. Lateral cavities are absent or indistinctly developed along the upper four-fifths of the lateral projection. A basal cavity is absent, but a shallow furrow can be observed in some specimens.
Materials.—
86 specimens.
Occurrence.—
Known from Wangcun and Wa’ergang sections, where it ranges from the Westergaardodina lui-Westergaardodina ani Zone through the Cordylodus lindstromi Zone (lower part).
Comparison.—
The present species differs from Westergaardodina ligula in its slender appearance and the different lateral cavities. Moreover, it appears considerably lower stratigraphically than W. ligula.
Remarks.—
The present species Westergaardodina ani was previously called Westergaardodina proligula, which is a nomen nudum (Dong, 1999, Reference Dong2000).
Westergaardodina cf. behrae Müller and Hinz, Reference Müller and Hinz1991
1991 cf. Westergaardodina behrae Reference Müller and HinzMüller and Hinz, p. 42, pl. 37, figs. 1–6, 9, 10, 12, 13.
Materials.—
12 specimens.
Occurrence.—
Known from the Wangcun section, where it occurs in the Westergaardodina quadrata Zone.
Remarks.—
The specimens at hand agree in gross morphology with Westergaardodina behrae but differ in its much larger size and the converging, shorter lateral projections.
Westergaardodina brevidens Dong, Reference Dong1993
1993Westergaardodina brevidens Reference DongDong, p. 353, pl. 1, figs. 6, 7.
2001aWestergaardodina brevidens Dong; Reference Dong and BergströmDong and Bergström, p. 973, pl. 1, fig. 8, pl. 2, fig. 8, pl. 6, fig 6.
Materials.—
28 specimens.
Occurrence.—
Known from Paibi and Wangcun sections, where it ranges from the Gapparodus bisulcatus-Westergaardodina brevidens Zone through the Shandongodus priscus-Hunanognathus tricuspidatus Zone.
Remarks.—
The present specimens agree well with those described and figured by Dong and Bergström (2001a).
Westergaardodina cf. calix Müller and Hinz, Reference Müller and Hinz1991
Figure 35.10–35.20
1991 cf. Westergaardodina cf. calix Reference Müller and HinzMüller and Hinz, p. 43, pl. 37, figs. 7, 8, 11, 14–16.
2004cWestergaardodina cf. calix Müller and Hinz; Reference Dong, Donoghue, Cheng and LiuDong et al., p. 1197, pl. 1, figs. 15–20, 22–25, 28.
Description.—
Small- to medium-size, bilaterally symmetrical, tricuspidate elements. The profile is rather flat. The median projection is narrower and longer than the lateral ones. In relation to the total length, the turning points are fairly high. The anterior side is smooth, with a wide central depression. The posterior side is a little smaller than the anterior one, especially along the lateral margin. The median projection bears a distinct rod-like keel, which extends above the turning points but does not reach the basal margin. Lateral cavities are developed in most specimens along the upper part of the lateral projections. Basal cavity is absent.
Materials.—
78 specimens.
Occurrence.—
Known from the Wangcun and Wa’ergang sections, where it ranges from the Westergaardodina cf. calix-Prooneotodus rotundatus Zone through the Cordylodus intermedius Zone.
Remarks.—
This species includes specimens which are not well preserved and previously were included in Westergaardodina cf. behrae. The present form resembles Westergaardodina calix in gross morphology, but it differs in its narrower and longer median projection, and in being thicker. Moreover, it appears lower stratigraphically than W. calix. Because the majority of the specimens of Westergaardodina cf. calix are not completely preserved in that one, two, or three of the upper parts of the three projections are missing, we do not propose it as a new species. The main difference between Westergaardodina behrae and W. cf. calix is in the height of the turning points, the width of the median projection, and the relative lengths of the elements (Müller and Hinz, Reference Müller and Hinz1991, p. 42).
Westergaardodina dimorpha new species
Figures 35.22–35.26, 36.1–36.4

Figure 36 (1–4) Westergaardodina dimorpha n. sp.; (1) Bitiao Formation, Wangcun section, GMPKU2982, posterior view; (2) Bitiao Formation, Wangcun section, GMPKU2983, posterior view; (3) Bitiao Formation, Wangcun section, GMPKU2984, anterior view; (4) Bitiao Formation, Wangcun section, GMPKU2985, posterior view; (5–9, 13) Westergaardodina elegans Dong and Bergström, 2001a; (5) Chefu Formation, Paibi section, GMPKU2986, posterior view; (6) Chefu Formation, Paibi section, GMPKU2987, posterior view; (7) Chefu Formation, Paibi section, GMPKU2988, anterior view; (8) Chefu Formation, Paibi section, GMPKU2989, posterior view; (9) Chefu Formation, Paibi section, GMPKU2990, posterior view; (13) Chefu Formation, Paibi section, GMPKU2991, posterior view; (10–12) Westergaardodina gigantea n. sp.; (10) Chefu Formation, Wangcun section, GMPKU2992, posterior view; (11) Chefu Formation, Wangcun section, GMPKU2993, posterior view; (12) Chefu Formation, Wangcun section, holotype, GMPKU2994, posterior view; (14–17) Westergaardodina grandidens Dong, Reference Dong1993; (14) Chefu Formation, Paibi section, GMPKU2995, posterior view; (15) Chefu Formation, Wangcun section, GMPKU2996, posterior view; (16) Chefu Formation, Wangcun section, GMPKU2997, posterior view; (17) Chefu Formation, Paibi section, large broken basal part, GMPKU2998, posterior view; (18, 19) Westergaardodina horizontalis Dong, Reference Dong1993; (18) Huaqiao Formation, Paibi section, GMPKU2999, posterior view; (19) Huaqiao Formation, Paibi section, GMPKU3000, posterior view; (20, 21) Westergaardodina cf. horizontalis Dong, Reference Dong1993; (20) Chefu Formation, Wangcun section, GMPKU3001, oblique posterior view; (21) Chefu Formation, Wangcun section, GMPKU3002, oblique posterior view; (22–25) Westergaardodina lui Dong, Repetski and Bergström, 2004c; (22) Bitiao Formation, Wangcun section, GMPKU2182, posterior view; (23) Bitiao Formation, Wangcun section, GMPKU2183, posterior view; (24) Shenjiawan Formation, Wangcun section, GMPKU3003, posterior view; (25) Shenjiawan Formation, Wangcun section, GMPKU3004, posterior view. Relative scale bar represents 156 μm (1, 2, 4, 7, 13–17, 19, 20, 24, 25), 145 μm (3, 5, 9, 18, 22, 23), 136 μm (6, 8), 218 μm (10–12), 199 μm (21).
Diagnosis.—
Small, asymmetrical, bicuspidate elements comprising left and right forms. One projection is much longer than the other. No basal cavity. The lateral cavities are very small or there are no lateral cavities.
Description.—
Small-sized, asymmetrical, bicuspidate elements comprising left and right forms. The lateral projections are not of equal size; one is much longer than the other. They are subcircular in the lower part of elements and diverge at a very small angle at first, but the angle increases upward and the projections are recurved outward. The anterior side is flat or a little convex, whereas the posterior side is concave. Both projections are fused up to a level that is much higher than the turning points or are fused up to the tips of the projections, so there are no basal cavities. The very small lateral cavities are restricted only to the upper parts of the projections or there are no lateral cavities. A broad bulge may be developed in the basal region. Sometimes, the longer projection is broken and the recurved upper part is not preserved. Left and right forms occur in about equal numbers.
Etymology.—
dimorpha, Latin, dimorphic.
Types.—
Holotype: GMPKU2980 from the Westergaardodina lui-Westergaardodina ani Zone, Bitiao Formation, Wangcun section, Wangcun village, Yongshun county, Hunan Province.
Materials.—
200 specimens
Occurrence.—
Known from the Wangcun section, where it occurs in the Westergaardodina lui-Westergaardodina ani Zone.
Westergaardodina elegans Dong and Bergström, 2001a
1993Westergaardodina sp. A. Reference DongDong, p. 355, pl. 2, figs. 1–4.
2001aWestergaardodina elegans Reference Dong and BergströmDong and Bergström, p. 973, pl. 3, fig. 18, pl. 4, figs. 9, 15, pl. 6, fig. 7.
2006Westergaardodina elegans Dong and Bergström; Reference Qi, Bagnoli and WangQi et al., p. 187, pl. 3, figs. 5, 7.
Materials.—
266 specimens.
Occurrence.—
Known from Paibi and Wangcun sections, where it ranges from the Shandongodus priscus-Hunanognathus tricuspidatus Zone through the Westergaardodina lui-Westergaardodina ani Zone.
Remarks.—
The present specimens agree well with those described and figured by Dong and Bergström (Reference Dong and Bergström2001a).
Westergaardodina gigantea new species
Diagnosis.—
Large, symmetrical, bicuspidate elements. The posterior side is much smaller than the anterior side. The turning point is located very high.
Description.—
Large-sized, symmetrical, bicuspidate elements. The anterior side is convex, whereas the posterior side is concave. The posterior side is much smaller than the anterior side. Both projections are fused at the bases. The turning points are located very high. The lateral projections are of equal size. Tips of projections are directed slightly outward, beginning near the tops of the projections. The large lateral openings wrap around the base and are exposed on the anterior sides of the base.
Etymology.—
gigantea, Latin, gigantic.
Types.—
Holotype: GMPKU2994 from the Westergaardodina matsushitai-Westergaardodina grandidens Zone, Chefu Formation, Wangcun section, Wangcun village, Yongshun county, Hunan Province.
Materials.—
12 specimens.
Occurrence.—
Known only from Wangcun section, where it occurs in the Westergaardodina matsushitai-Westergaardodina grandidens Zone.
Remarks.—
Westergaardodina gigantea n. sp. is distinguished by its large size and by the turning points being located very high.
Westergaardodina grandidens Dong, Reference Dong1993
1978Westergaardodina moessebergensis (Müller); Reference AbaimovaAbaimova, p. 503, pl. 8, figs. 8, 10, ?11.
1991Westergaardodina matsushita (Nogami); Reference Müller and HinzMüller and Hinz, pl. 28, fig. 17 only (not figs. 15–16, 18–20). (part)
1993Westergaardodina grandidens Reference DongDong, p. 354, pl. 3, figs. 5, 8, 9.
2001aWestergaardodina grandidens Dong; Reference Dong and BergströmDong and Bergström, p. 973, pl. 4, figs. 1, 5, 7.
2013Westergaardodina grandidens Dong; Reference LeeLee, p. 169, pl. 2, figs. 10, 23–27.
Materials.—
66 specimens.
Occurrence.—
known from the Paibi and Wangcun sections, where it occurs in the Westergaardodina matsushitai-Westergaardodina grandidens Zone.
Remarks.—
The present specimens agree well with those described and figured by Dong and Bergström (Reference Dong and Bergström2001a).
Westergaardodina horizontalis Dong, Reference Dong1993
1993Westergaardodina horizontalis Reference DongDong, p. 354, pl. 1, figs. 13–14.
2001aWestergaardodina horizontalis Dong; Reference Dong and BergströmDong and Bergström, p. 974, pl. 6, figs. 13–14.
? 2008Westergaardodina horizontalis Dong; Reference Bagnoli, Qi and WangBagnoli et al., p. 112, fig. 3 (M).
Materials.—
12 specimens.
Occurrence.—
Known from the Paibi and Wangcun sections, where it occurs in the Gapparodus bisulcatus-Westergaardodina brevidens Zone.
Remarks.—
The present specimens agree well with those described and figured by Dong and Bergström (Reference Dong and Bergström2001a).
Westergaardodina cf. horizontalis Dong, Reference Dong1993
cf. 1993 Westergaardodina horizontalis Reference DongDong, p. 354, pl. 1, figs. 13–14.
2001aWestergaardodina cf. horizontalis Dong; Reference Dong and BergströmDong and Bergström, p. 976, pl. 2, fig. 21.
Materials.—
Six specimens.
Occurrence.—
Known from the Wangcun secton, where it occurs in the Westergaardodina matsushitai-Westergaardodina grandidens Zone.
Remarks.—
This form is similar to Westergaardodina horizontalis in having two horizontally extending, wing-like lateral projections, but the keel on the median projection is much shorter than in W. horizontalis s. s., and it is solid rather than hollow. The keel extends from the tip to a point approximately one-third of the length of the median projection, where the posterior side of the element is nearly perpendicular to the flat anterior side, forming deep and broad basal and lateral cavities.
Westergaardodina lui Dong, Repetski, and Bergström, 2004c
Figures 36.22–36.25, 37.1–37.3, 37.7–37.9

Figure 37 (1–3, 7–9) Westergaardodina lui Dong, Repetski, and Bergström, 2004c; (1) Bitiao Formation, Wangcun section, GMPKU3005, posterior view; (2) Bitiao Formation, Wangcun section, GMPKU3006, posterior view; (3) Bitiao Formation, Wangcun section, GMPKU3007, anterior view; (7) Bitiao Formation, Wangcun section, GMPKU3008, posterior view; (8) Bitiao Formation, Wangcun section, GMPKU3009, posterior view; (9) Bitiao Formation, Wangcun section, GMPKU3010, posterior view; (4–6, 10–12) Westergaardodina matsushitai Nogami, Reference Nogami1966; (4) Chefu Formation, Wangcun section, GMPKU3011, posterior view; (5) Chefu Formation, Wangcun section, GMPKU3012, posterior view; (6) Chefu Formation, Wangcun section, GMPKU3013, posterior view; (10) Chefu Formation, Wangcun section, GMPKU3014, posterior view; (11) Chefu Formation, Wangcun section, GMPKU3015, posterior view; (12) Bitiao Formation, Wangcun section, GMPKU3016, posterior view; (13) (14, 16) Westergaardodina microdentata Zhang, 1983; (13) Bitiao Formation, Wangcun section, GMPKU3017, posterior view; (14) Bitiao Formation, Wangcun section, GMPKU3018, posterior view; (16) Bitiao Formation, Wangcun section, GMPKU3019, posterior view; (15) Westergaardodina cf. nogamii Müller and Hinz, Reference Müller and Hinz1991, Shenjiawan Formation, Wa’ergang section, GMPKU3020, posterior view; (17) Westergaardodina sp. E, Bitiao Formation, Wangcun section, GMPKU3021, posterior view; (18–26) Westergaardodina quadrata (An, Reference An1982); (18) Chefu Formation, Paibi section, GMPKU3022, posterior view; (19) Chefu Formation, Paibi section, GMPKU3023, posterior view; (20) Chefu Formation, Paibi section, GMPKU3024, posterior view; (21) Chefu Formation, Paibi section, GMPKU3025, posterior view; (22) Chefu Formation, Paibi section, GMPKU3026, posterior view; (23) Chefu Formation, Wangcun section, GMPKU3027, anterior view; (24) Chefu Formation, Wangcun section, GMPKU3028, posterior view; (25) Chefu Formation, Wangcun section, GMPKU3029, posterior view; (26) Chefu Formation, Wangcun section, GMPKU3030, posterior view. Relative scale bar represents 143 μm (1–3, 5, 6, 9–17, 19, 23–26), 136 μm (4, 7, 8, 18, 20, 21), 219 μm (22).
2004cWestergaardodina lui Dong, Repetski, and Bergström, p. 1197, pl. 3. figs. 4–10, 18–20, 22–24, 27.
Description.—
A small, bilaterally asymmetrical bicuspidate Westergaardodina. One lateral projection is longer than another, and there are right and left forms. The anterior side is strongly convex and is characterized by a deeply incised turning point. The posterior side is deeply concave, with the maximum depth at the turning point. Along the longer lateral projection, the posterior side is smaller than the anterior one, resulting in a shallow lateral cavity extending along most of the projection. The lateral cavity is missing or weakly developed within the upper one-third of the shorter lateral projection. The base is rather narrow.
Materials.—
98 specimens.
Occurrence.—
Known from the Wangcun and Wa’ergang sections, where it ranges from the Westergaardodina lui-Westergaardodina ani Zone through the Cordylodus intermedius Zone.
Comparison.—
The present species differs from Westergaardodina ligula in its narrow base and asymmetrical bicuspidate elements. It differs from Westergaardodina wimani in its narrow base, strongly convex anterior side, and deeply concave posterior side. Also, the lateral projections of W. wimani differ considerably in length.
Westergaardodina matsushitai Nogami, Reference Nogami1966
1966Westergaardodina matsushitai Reference NogamiNogami, p. 360, pl. 10, figs. 6–8.
1971Westergaardodina matsushitai Nogami; Reference MüllerMüller, fig. 1d.
1978Westergaardodina matsushitai Nogami; Reference AbaimovaAbaimova, p. 502, pl. 8, fig. 6.
1981Westergaardodina matsushitai Nogami; Reference AnAn, p. 219, pl. 1, fig. 4.
1982Westergaardodina matsushitai Nogami; Reference AnAn, p. 153, pl. 6, figs. 1–4.
1983Westergaardodina matsushitai Nogami; Reference An, Zhang, Xiang, Zhang, Xu, Zhang, Jiang, Yang, Lin, Cui and YangAn et al., p. 163, pl. 1, figs. 3, 4, 8.
1991Westergaardodina matsushitai Nogami; Reference Müller and HinzMüller and Hinz, p. 46, pl. 28, figs. 15, 16, 18–20 (not fig. 17). (part)
1993Westergaardodina matsushitai Nogami; Reference DongDong, p. 354, pl. 1, figs. 4, 5.
2001aWestergaardodina matsushitai Nogami; Reference Dong and BergströmDong and Bergström, p. 976, pl. 6, figs. 10–12.
? 2002a Westergaardodina matsushitai Nogami; Reference LeeLee, p. 164, pl. 1, fig. 1.
? 2002b Westergaardodina matsushitai Nogami; Reference LeeLee, p. 25, pl. 1, fig. 3.
2006Westergaardodina matsushitai Nogami; Reference Qi, Bagnoli and WangQi et al., p. 187, pl. 3, fig. 4.
Materials.—
47 specimens.
Occurrence.—
Known from the Paibi and Wangcun sections, where it occurs in the Westergaardodirna matsushitai-Westergaardodirna grandidens Zone.
Remarks.—
The specimens at hand agree well with those described and illustrated by Nogami (Reference Nogami1966) and Müller and Hinz (Reference Müller and Hinz1991), except that their plate 8, figure 7 is assigned herein to Westergaardodina grandidens. Some specimens exhibit a smaller than typical angle of divergence between the lateral projections, but that is considered to represent intraspecific variation. Müller and Hinz’s (Reference Müller and Hinz1998, p. 105) experimented on elements of Westergaardidina polymorpha and concluded that the angle of divergence is not of taxonomic value. However, that is not known whether or not this conclusion can be applied to other species of Westergaardodina.
Westergaardodina microdentata Zhang in An et al., Reference An, Zhang, Xiang, Zhang, Xu, Zhang, Jiang, Yang, Lin, Cui and Yang1983
1983Westergaardodina microdentata Zhang in Reference An, Zhang, Xiang, Zhang, Xu, Zhang, Jiang, Yang, Lin, Cui and YangAn et al., p. 164, pl. 1, figs. 15–17; text-fig. 9 (7).
1991Westergaardodina microdentata Zhang; Reference Müller and HinzMüller and Hinz, p. 47, pl. 40, figs. 8–15.
Materials.—
Nine specimens.
Occurrence.—
Known from the Wangcun section, where it occurs from the Westergaardodina lui-Westergaardodina ani Zone through the Westergaardodina cf. calix-Prooneotodus rotundatus Zone.
Remarks.—
The present specimens agree in all aspects with the previous descriptions and illustrations of Westergaardodina microdentata.
Westergaardodina cf. nogamii Müller and Hinz, Reference Müller and Hinz1991
cf. 1991 Westergaardodina nogamii Reference Müller and HinzMüller and Hinz, p. 47, pl. 39, figs. 1–10.
Materials.—
Only one specimen.
Occurrence.—
Known from the Wa’ergang section, where it occurs in the Eoconodontus Zone.
Remarks.—
The specimen at hand approximately agrees in gross morphology with Westergaardodina nogamii, but it differs in its much smaller size, diverging lateral projections, and shorter median projection. Nevertheless, the present specimen is insufficient to name a new species.
Westergaardodina quadrata (An, Reference An1982)
1982Westergaardodina moessebergensis quadrata Reference AnAn, p. 153, pl. 6, figs. 5–8, 10.
1983Westergaardodina moessebergensis (An), Reference An, Zhang, Xiang, Zhang, Xu, Zhang, Jiang, Yang, Lin, Cui and YangAn et al., p. 164, pl. 1, fig. 6 only.
1987Westergaardodina bicuspidata (Müller); Reference AnAn, p. 115, pl. 3, figs. 13, 18.
1991Westergaardodina quadrata (An); Reference Müller and HinzMüller and Hinz, p. 50, pl. 30, figs. 9, 11–21.
1993Westergaardodina quadrata (An); Reference DongDong, pl. 2, figs. 6, 7, 9, 10, 12.
2001aWestergaardodina quadrata (An); Reference Dong and BergströmDong and Bergström, p. 978, pl. 1, fig. 14, pl. 3, fig. 15, pl. 6, figs. 4, 8.
2006Westergaardodina quadrata (An); Reference Qi, Bagnoli and WangQi et al., p. 187, pl. 3, fig. 1.
2009Westergaardodina quadrata (An); Reference Lee, Lee, Miller and JeongLee et al., p. 425, fig. 8 (19), p. 427, fig. 9 (16).
Materials.—
79 specimens.
Occurrence.—
Known from the Paibi and Wangcun sections, where it ranges from the Westergaardodina quadrata Zone through the Westergaardodina matsushitai-Westergaardodina grandidens Zone.
Remarks.—
The specimens at hand agree well with those described and illustrated by An (Reference An1982) and Müller and Hinz (Reference Müller and Hinz1991).
Westergaardodina sola new species
2001aWestergaardodina sp. Reference Dong and BergströmDong and Bergström, p. 967, pl. 2, fig. 18.
Diagnosis.—
Medium-sized elements with unique outline. The posterior side is smaller than the anterior, with a prominent bulge on the posterior side. Both sides are fused along the upper margin, so that there is only one “projection.” Basal cavity is large and connected with the “lateral cavity”.
Description.—
Element is medium-sized, with an outline that is unique among all known species of Westergaardodina. The anterior side is flat, whereas the posterior side is concave with a prominent bulge. The posterior side is smaller than the anterior. Both sides are fused along the upper margin, which is formed into a sharp costa. Both sides extend upward on the left part of the element, leading to the formation of a structure similar to a projection. Basal cavity is large and connected with the opening of the projection-like structure. The size of the posterior side and the length and shape of the “lateral projection” are variable.
Etymology.—
sola, Latin, sole, referring to the unique outline of the element.
Types.—
Holotype: GMPKU3033 from the Westergaardodina quadrata Zone, Chefu Formation, Paibi section, Paibi village, Huayuan county, Hunan Province.
Materials.—
Nine specimens.
Occurrence.—
Known from the Wangcun and Paibi sections, where it ranges from Westergaardodina quadrata Zone through the Westergaardodina matsushitai-Westergaardodina grandidens Zone.
Westergaardodina tetragonia Dong, Reference Dong1993
1993Westergaardodina tetragonia Reference DongDong, p. 355, pl. 1, figs. 1–3, 9.
2001aWestergaardodina tetragonia Dong; Reference Dong and BergströmDong and Bergström, p. 979, pl. 6, figs. 1, 3.
non2006 Westergaardodina tetragonia Dong; Reference Qi, Bagnoli and WangQi et al., p. 187, pl. 3, fig. 6.
Materials.—
45 specimens.
Occurrence.—
Known from the Paibi and Wangcun sections, where it ranges from the Shandongodus priscus-Hunanognathus tricuspidatus Zone through the Westergaardodina quadrata Zone.
Remarks.—
The present specimens agree well with those described and figured by Dong and Bergström (Reference Dong and Bergström2001a).
Westergaardodina tricuspidata Müller, Reference Müller1959
1959Westergaardodina tricuspidata Reference MüllerMüller, p. 470, pl. 15, figs. 5, 6.
1965Westergaardodina tricuspidata Müller; Reference GrantGrant, p. 145, pl. 15, fig. 31.
1979Westergaardodina tricuspidata Müller; Reference BednarczykBednarczyk, p. 436, pl. 2, fig. 5.
1991Westergaardodina tricuspidata Müller; Reference Müller and HinzMüller and Hinz, p. 50, pl. 36, figs. 1–4, 6.
Materials.—
100 specimens.
Occurrence.—
Known from the Wangcun and Wa’ergang sections, where it ranges from the Westergaardodina lui-Westergaardodina ani Zone through the Westergaardodina cf. calix-Prooneotodus rotundatus Zone.
Remarks.—
The specimens at hand agree well with those described and illustrated by Müller (Reference Müller1959) and Müller and Hinz (Reference Müller and Hinz1991).
Westergaardodina wimani Szaniawski, Reference Szaniawski1971
1971Westergaardodina wimani Reference SzaniawskiSzaniawski, p. 409, pl. 1, fig. 5, pl. 5, figs. 7–9.
1982Westergaardodina wimani Szaniawski; Reference AnAn, p. 156, pl. 9, fig. 9 only (not fig. 14). (part)
1991Westergaardodina wimani Szaniawski; Reference Müller and HinzMüller and Hinz, p. 51, pl. 28, figs. 21–27.
2004cWestergaardodina wimani Szaniawski; Reference Dong, Donoghue, Cheng and LiuDong et al., p. 1205, pl. 3, fig. 29.
Materials.—
Eight specimens.
Occurrence.—
Known from the Wangcun section, where it ranges from the Westergaardodina cf. calix-Prooneotodus rotundatus Zone through the Proconodontus Zone.
Remarks.—
The present specimens agree well with those described and illustrated by Szaniawski (Reference Szaniawski1971) and Müller and Hinz (Reference Müller and Hinz1991). Müller and Hinz (Reference Müller and Hinz1991) assigned Westergaardodina semitricuspidata An, Reference An1982 to Westergaardodina wimani. However, An (Reference An1982) erected his species based on only two broken specimens. In the light of additional similar specimens recovered from Hunan, the characters of this species are much more evident.
Westergaardodina sp. A
Description.—
Large-sized, symmetrical, bicuspidate element. The turning point is located very high. The anterior side is flat, whereas the posterior side is slightly concave. The lateral projections extend from the triangular lower part of the element and diverge greatly, leading to an anchor-shaped profile of the element. Large lateral cavities extend from tips to bases of projections and connect with the basal cavity.
Materials.—
Only one specimen.
Occurrence.—
Known from Wangcun section, where it occurs in the Westergaardodina cf. calix-Prooneotodus rotundatus Zone.
Remarks.—
Westergaardodina sp. A is distinguished by its large size, very high turning point, and the anchor-shaped profile. Nevertheless, the present specimen is inadequate to name a new species.
Westergaardodina sp. B
Description.—
Large-sized, symmetrical, bicuspidate element. The turning point is located relatively high. The anterior side is slightly convex, whereas the posterior side is concave. Both lateral projections are fused up to a level that is above the turning point. The lateral projections extend from the semicircular lower part of element and diverge slightly. Tips of projections directed slightly outward, and lateral openings are only in the upper parts of the projections. The lower part of element is very thick antero-posteriorly.
Materials.—
Only one specimen.
Occurrence.—
Known from the Wa’ergang sections, where it occurs in the Westergaardodina lui-Westergaardodina ani Zone.
Remarks.—
Westergaardodina sp. B is distinguished by its large size, relatively high turning point, and antero-posteriorly thick, semicircular lower part of element. The present specimen is insufficient to name a new species.
Westergaardodina sp. C
Description.—
Large subsymmetrical, tricuspidate element. The anterior side is convex, whereas the posterior side is concave. The median projection is smaller than the lateral ones. It is characterized by a long keel which extends from tip to base and ends with a conical opening. The tips of lateral projections are directed outward. The large basal cavity is continuously open near the base.
Materials.—
Eight specimens, but only one specimen is well preserved.
Occurrence.—
Known from the Wa’ergang sections, where it occurs in the Eoconodontus Zone.
Comparison.—
The specimen at hand are somewhat similar to Westergaardodina amplicava Müller, Reference Müller1959 in gross morphology. They differ from the latter in having a wider median projection that has a larger funnel-shaped keel. Also, there are no keels on the lateral projections of the present species.
Westergaardodina sp. D
Description.—
Small-sized, symmetrical, bicuspidate elements. The lateral projections diverge at an acute angle. The turning points are located relatively high. The anterior side is convex, whereas the posterior side is concave. Both vertical projections are fused up to a level that is a little below the turning points. The lateral cavities are small and extend upward to just a little below the turning points. A broad bulge is developed in the basal region.
Materials.—
Nine specimens, but only one specimen is well preserved.
Occurrence.—
Known from the Wangcun section, where it occurs in the Shandongodus priscus-Hunanognathus tricuspidatus Zone.
Westergaardodina sp. E
Description.—
Medium-sized, subsymmetrical, bicuspidate elements which are distinguished by the broad U-shaped or saddle-shaped profile. There is no conspicuous boundary between the base and the two lateral projections, which extend laterally. The anterior side is convex, whereas the posterior side is concave and smaller than the anterior side. Both sides are fused along the entire upper margin of the element. Basal cavity is shallow and connected with the lateral cavities in some specimens that have deep lateral cavities.
Materials.—
Eight specimens, but only one specimen is well preserved.
Occurrence.—
Known from Wangcun section, where it occurs in the Westergaardodina cf. calix-Prooneotodus rotundatus Zone.
Comparison.—
Westergaardodina sp. E differs from any konwn species of Westergaardodina in its peculiar profile.
Genus Yongshunella Dong and Bergström, Reference Dong and Bergström2001a
Type species.—
Yongshunella polymorpha Dong and Bergström, Reference Dong and Bergström2001a.
Yongshunella polymorpha Dong and Bergström, Reference Dong and Bergström2001a
1993Hertzina? sp. A. Reference DongDong, p. 351, pl. 5, figs. 1, 2, 6.
2001aYongshunella polymorpha Reference Dong and BergströmDong and Bergström, p. 980, pl. 1, fig. 20, pl. 2, figs. 1–7, 14, 22, pl. 4, figs. 8, 14.
2006Yongshunella polymorpha Dong and Bergström; Reference Qi, Bagnoli and WangQi et al., p. 187, pl. 3, fig. 16.
2008Yongshunella polymorpha Dong and Bergström; Reference Bagnoli, Qi and WangBagnoli et al., p. 112, Fig. 3 (C–E).
Materials.—
120 specimens.
Occurrence.—
Known from the Paibi and Wangcun sections, where it ranges from the Gapparodus bisulcatus-Westergaardodina brevidens Zone through the Westergaardodina lui-Westergaardodina ani Zone.
Remarks.—
The present specimens agree well with described and figured by Dong and Bergström (Reference Dong and Bergström2001a).
Protoconodonts
Phylum Chaetognatha Leuckart, Reference Leuckart1854
Order Protoconodontida Landing, Reference Landing1995
Genus Gapparodus Abaimova, Reference Abaimova1978
Type species.—
Hertzina? bisulcatus Müller, Reference Müller1959.
Gapparodus bisulcatus (Müller, Reference Müller1959)
1959Hertzina? bisulcatus Reference MüllerMüller, p. 456, pl. 13, figs. 22, 23, 27.
1966Hertzina? bisulcatus Müller; Reference PoulsenPoulsen, p. 8, pl. 1, fig. 9, text-fig. 3.
1969Hertzina bisulcatus Müller; Reference Clark and RobisonClark and Robison, p. 1045, fig. 1 (d).
1969Hertzina bisulcatus Müller; Reference Clark and MillerClark and Miller, fig. 1 (2–4).
1971Hertzina bisulcatus Müller; Reference MüllerMüller, p. 12, pl. 1, fig. 7.
1973Hertzina bisulcatus Müller; Reference Özgül and GedikÖzgül and Gedik, p. 47, pl. 1, fig. 8.
1974Hertzina bokononi Reference LandingLanding, p. 1246, text-fig. 1 (h, i).
1974Hertzina bisulcatus Müller; Reference LandingLanding, p. 1246, text-fig. 1 (c, d, e, f). (with var. 1 and 2)
1975Hertzina bisulcatus Müller; Reference LeeLee, p. 80, pl. 1, figs. 2c, 3.
1976Hertzina? bisulcatus Müller; Reference BengtsonBengtson, p. 191, figs. 5–9.
1976Hertzina? bisulcatus Müller; Reference Abaimova and ErgalievAbaimova and Ergaliev, p. 392, pl. 14, figs. 12–15.
1978Gapparodus heckeri Reference AbaimovaAbaimova, p. 496, pl. 7, figs. 3, 4, text-fig. 1 (e).
1978Gapparodus bellus Reference AbaimovaAbaimova, p. 497, pl. 7, figs. 6, 7, text-fig. 1 (g).
1978Gapparodus bisulcatus (Müller); Reference AbaimovaAbaimova, p. 497, p. 7, fig. 8, text-fig. 1 (h).
1978Gapparodus porrectus Reference AbaimovaAbaimova, p. 497, p. 7, fig. 5, text-fig. 1 (i).
1987Gapparodus bisulcatus (Müller); Reference AnAn, p. 106, pl. 1, figs. 6, 8–9, 26. (part)
1990Gapparodus bisulcatus (Müller); Reference An and ZhengAn and Zheng, p. 159, pl. 2, figs. 2, 8.
1991Gapparodus bisulcatus (Müller); Reference Müller and HinzMüller and Hinz, p. 25, pl. 3, figs. 1–42.
1993Gapparodus bisulcatus (Müller); Reference DongDong, pl. 6, figs. 1, 3, 9, 12.
1994Gapparodus bisulcatus (Müller); Reference An and MeiAn and Mei, pl. 1, fig. 30.
1998Gapparodus bisulcatus (Müller); Reference Müller and HinzMüller and Hinz, p. 96, fig. 2 (1–11).
2001aGapparodus bisulcatus (Müller); Reference Dong and BergströmDong and Bergström, p. 964, pl. 3, figs. 1, 3, pl. 5, fig. 11.
2006Gapparodus bisulcatus (Müller); Reference Qi, Bagnoli and WangQi et al., p. 187, pl. 3, fig. 15.
2008Gapparodus bisulcatus (Müller); Reference Bagnoli, Qi and WangBagnoli et al., p. 112, fig. 3 (O).
2009Gapparodus bisulcatus (Müller); Reference Lee, Lee, Miller and JeongLee et al., p. 420, fig. 6 (1, 11, 15, 22).
Description.—
Moderate to large, slender coniform elements, evenly recurved over their entire length, so there is no conspicuous junction between cusp and base. Both anterior and posterior sides are convex. The lateral sides are characterized by distinct furrows, the position of which is variable. Basal cavities are very deep, with the apex extending nearly to the tip of the element. The cross section is circular at the apical portion and changes into a variable outline toward the base.
Materials.—
Approximately 1200 specimens.
Occurrence.—
Known from the Paibi, Wangcun and Wa’ergang sections, where it ranges from the Gapparodus bisulcatus-Westergaardodina brevidens Zone through the Westergaardodina lui-Westergaardodina ani Zone. The species is common in South China, Sweden, Denmark, and Kazakhstan and it has also been found in Ordos, northwestern China and eastern New York State. It is rare in most of North China and in most of North America. Its distribution suggests that it most probably preferred cold-water environments.
Remarks.—
The specimens at hand agree in all respects with the elements described by Müller and Hinz (Reference Müller and Hinz1991).
Genus Gumella Müller and Hinz, Reference Müller and Hinz1991
Type species.—
Gumella cuneata Müller and Hinz, Reference Müller and Hinz1991.
Gumella cuneata Müller and Hinz, Reference Müller and Hinz1991
1971Hertzina elongata Reference MüllerMüller, pl. 1, fig. 2.
1987Gapparodus bisulcatus (Müller); Reference AnAn, p. 106, pl. 1, fig. 10. (part)
1991Gumella cuneata Reference Müller and HinzMüller and Hinz, p. 26, pl. 4, figs. 1–25, pl. 5, figs. 1–28.
1998Gumella cuneata Müller and Hinz; Reference Müller and HinzMüller and Hinz p. 97, figs. 3, 4 (1–7), 5 (1–5).
2001aGumella cuneata Müller and Hinz; Reference Dong and BergströmDong and Bergström, p. 964, pl. 1, figs. 5–7, 11, 19.
2008Gumella cuneata Müller and Hinz; Reference Bagnoli, Qi and WangBagnoli et al., p. 112, fig. 3 (N).
2009Gumella cuneata Müller and Hinz; Reference Lee, Lee, Miller and JeongLee et al., p. 421, fig. 6 (19–21), p. 425, fig. 8 (15).
Description.—
Coniform elements, moderate to large, proclined, with no conspicuous junction between cusp and base. Anterior sides convex, posterior sides flattened. Lateral sides characterized by symmetrical posterolateral furrows, the posterior sides of which are reinforced by an additional unit. Basal cavity very large and deep, with its apex extending nearly to the tip of the element. The cross section is circular at the apical portion and it changes into a variable outline toward the base.
Materials.—
280 specimens.
Occurrence.—
Known from the Paibi, Wangcun, and Wa’ergang sections, where it ranges from the Gapparodus bisulcatus-Westergaardodina brevidens Zone through the Westergaardodina lui-Westergaardodina ani Zone.
Remarks.—
Gumella cuneata and Gapparodus bisulcatus are similar in gross morphology, but the former species has more distinct and deeper lateral furrows reinforced by an additional unit (Müller and Hinz, Reference Müller and Hinz1991, p. 26). Although the internal structure of the two species represents the same type, the structure of G. cuneata appears to be somewhat more complex. The fact that elements of G. cuneata and G. bisulcatus frequently occur together and are quite similar in external and internal morphology may indicate that they represent the same multi-element species.
Genus Huayuanodontus Dong and Bergström, 2001a
Type species.—
Albiconus? tricornis Dong, Reference Dong1993.
Huayuanodontus tricornis (Dong, Reference Dong1993)
Figure 19.14–19.16, 19.22–19.24
1993Albiconus? tricornis Reference DongDong, p. 349, pl. 6, figs. 4, 5, 11, 15, 20–22.
2001aHuayuanodontus tricornis (Dong); Reference Dong and BergströmDong and Bergström, p. 964, pl. 2, figs. 15–16, pl. 3, figs. 2, 4, 8, 11.
2006Huayuanodontus tricornis (Dong); Reference Qi, Bagnoli and WangQi et al., p. 187, pl. 3, figs. 13–14.
2008Huayuanodontus tricornis (Dong); Reference Bagnoli, Qi and WangBagnoli et al., p. 112, fig. 3 (I).
2013Huayuanodontus tricornis (Dong); Reference LeeLee, p. 168, pl. 1, figs. 24–26.
Materials.—
600 specimens.
Occurrence.—
Present in the Paibi, Wangcun, and Wa’ergang sections, where it ranges from the Gapparodus bisulcatus-Westergaardodina brevidens Zone through the Proconodontus tenuiserratus Zone.
Remarks.—
The specimens at hand agree well with Huayuanodontus tricornis described and figured by Dong and Bergström (Reference Dong and Bergström2001a). The present species differs from Albiconus postcostatus Miller in having a triangular, rather than roundly triangular, cross-section, and a more slender shape of the element which is not laterally compressed, as is Miller’s species. The present species occurs in significantly older strata than A. postcostatus, which is present in strata near the Cambrian–Ordovician boundary. The present form looks like a protoconodont. However, preliminary histological investigation indicates it is neither a protoconodont nor a paraconodont. It is also not a euconodont (Dong, Reference Dong2007a). Therefore, further detailed histological work is needed to substantiate the nature of the present genus, although it seems beyond the scope of this study.
Genus Paibiconus Dong, Reference Dong1993
Type species.—
Paibiconus proarcuatus Dong, Reference Dong1993.
Paibiconus proarcuatus Dong, Reference Dong1993
1993Paibiconus proarcuatus Reference DongDong, p. 353, pl. 5, figs. 12–16.
2001aPaibiconus proarcuatus Dong; Reference Dong and BergströmDong and Bergström, p. 969, pl. 1, fig. 4, pl. 2, fig. 11, pl. 3, figs. 5–7.
2008Paibiconus proarcuatus Dong; Reference Bagnoli, Qi and WangBagnoli et al., p. 112, fig. 3 (Q).
Materials.—
660 specimens.
Occurrence.—
Known from the Paibi, Wa’ergang and Wangcun sections, where it occurs from the Gapparodus bisulcatus-Westergaardodina brevidens Zone through the Westergaardodina matsushitai-Westergaardodina grandidens Zone.
Remarks.—
The specimens at hand agree well with those described and figured by Dong and Bergström (Reference Dong and Bergström2001a). Histological investigation verified that this form is a protoconodont (Dong, Reference Dong2004).
Genus Phakelodus Miller, Reference Miller1984
Type species.—
Oneotodus tenuis (Müller, Reference Müller1959).
Remarks.—
Müller and Hinz (Reference Müller and Hinz1991, p. 32) separated specimens of Oneotodus tenuis into two taxa, Phakelodus tenuis and Phakelodus elongatus, stating “both P. elongatus and P. tenuis are long-ranging and widespread”. Although they appear in strikingly similar clusters, they were not regarded as morphotypes of a single species. All recognized clusters are unimembrate. Further, the occurrence of these species is indeed overlapping but not consistent enough (Müller and Hinz, Reference Müller and Hinz1991, table 3) for them to represent different morphotypes of a single species.
We have numerous specimens of these taxa from Hunan, where both are long-ranging and co-occurring, and are found unimemberate clusters that contain either the Phakelodus tenuis type or the Phakelodus elongatus type. However, this fact does not prove that these forms represent different species because it is quite possible that a protoconodont animal could have possessed more than one type of cluster. It is also possible that this could be an example of sexual dimorphism. In this study, we consider both these two types of elements, P. tenuis type (morphotype A) and P. elongatus type (morphotype B) to represent P. tenuis.
No clusters of the stout species Phakelodus simplex Müller and Hinz, Reference Müller and Hinz1991 have been found anywhere. In gross morphology, that species is more reminiscent of the paraconodont Prooneotodus gallatini, except that P. simplex has a protoconodont internal structure. It may be that P. simplex should not be referred to Phakelodus.
Phakelodus tenuis (Müller, Reference Müller1959)
1959Oneotodus tenuis Reference MüllerMüller, p. 457, pl. 13, figs. 11, 13, 14, 20.
1990Prooneotodus tenuis (Müller); Reference An and ZhengAn and Zheng, p. 160, pl. 1, figs. 1, 6, 7.
1991Phakelodus tenuis (Müller); Reference Müller and HinzMüller and Hinz, p. 33, pl. 1, figs. 6, 10, 11, 15–21, 23, pl. 2, figs. 1–24. (including extensive synonymy to 1988).
1991Phakelodus elongatus (An); Reference Müller and HinzMüller and Hinz, p. 32, pl. 1, figs. 1–5, 7–9, 12–14, 22. (including extensive synonymy to 1988).
1993Phakelodus tenuis (Müller); Reference DongDong, pl. 4, figs. 12, 13, pl. 6, figs. 2, 8, 17.
1993Phakelodus tenuis (Müller); Reference LandingLanding, p. 6, fig. 4 (1).
1993Phakelodus elongatus (An); Reference DongDong, pl. 4, figs. 10, 11, pl. 6, figs. 6, 7.
1994Phakelodus tenuis (Müller); Reference An and MeiAn and Mei, pl. 1, fig. 23.
1998Phakelodus elongatus (An); Reference Rao and TortelloRao and Tortello, p. 45, pl. 2, figs. 16, 17.
1999Phakelodus tenuis (Müller); Göncüoğlu and Kozur, p. 599, fig. 4 (B).
2000Phakelodus tenuis (Müller); Reference Pyle and BarnesDubinina, p. 166, pl. 1, fig. 7, pl. 2, figs. 10–12, 25.
2000Phakelodus? cavatua Reference Pyle and BarnesDubinina, p. 166, pl. 1, figs. 19, 19a, 27, 27a.
2001aPhakelodus tenuis (Müller); Reference Dong and BergströmDong and Bergström, p. 970, pl. 3, fig. 19, pl. 5, figs. 8, 10, pl. 6, fig. 5.
2001Phakelodus tenuis (Müller); Reference LeeLee, p. 449, fig. 6 (1).
2002Phakelodus elongatus (An); Lee, p. 25, pl. 1, fig. 1.
2002Phakelodus tenuis (Müller); Lee, p. 25, pl. 1, fig. 2.
2002Phakelodus tenuis (Müller); Lee, p. 164, pl. 1, fig. 8.
2004Phakelodus elongatus (An); Reference LeeLee, p. 60, fig. 5 (A).
2007‘Phakelodus elongatus’ (Zhang in An et al., Reference An, Zhang, Xiang, Zhang, Xu, Zhang, Jiang, Yang, Lin, Cui and Yang1983); Reference Landing, Westrop and KeppieLanding et al., fig. 6 (u).
2007Phakelodus tenuis (Müller); Reference Landing, Westrop and KeppieLanding et al., fig. 6 (s).
2008Phakelodus? sp. Reference Bagnoli, Qi and WangBagnoli et al., p. 112, fig. 3 (H).
2009Phakelodus tenuis (Müller); Reference Lee, Lee, Miller and JeongLee et al., p. 420, fig. 6 (12, 14).
2009Phakelodus elongatus (An); Reference Lee, Lee, Miller and JeongLee et al., p. 420, fig. 6 (9, 10).
Materials.—
Approximately 20,000 specimens.
Occurrence.—
Phakelodus tenuis is present in the Paibi, Wangcun, and Wa’ergang sections. It ranges from the Gapparodus bisulcatus-Westergaardodina brevidens Zone through the Cordylodus lindstromi Zone (Lower Part).
Remarks.—
Our specimens agree well with Phakelodus tenuis type (morphotype A) and Phakelodus elongatus type (morphotype B), respectively.
Acknowledgments
X.-P Dong thanks W.-C Lai and W. Guo for field assistance; S. M. Bergström, J. E. Repetski, and S. Stouge for their suggestions and comments; and P.C.J. Donoghue for his work on conodont histology and for his valuable advice on this manuscript. This manuscript is greatly improved by the substantive scientific and editorial comments of the reviewers J.E. Repetski and J.F. Miller. This work was supported by the National Natural Science Foundation of China (grant nos. 41372015, 41072006, 40772008, 40572003, 40372001, 40072007, 49772083, and 49272077 to X.P.D.; 41102003 and J1210006 to H.Z.), State Key Laboratory of Palaeobiology and Stratigraphy, Nanjing Institute of Geology and Palaeontology, Chinese Academy of Sciences (grant nos. 103102, 083101, 023106, 973103, and 913114 to X.P.D.; 20132107 to H.Z.), the Research Fund for Doctoral Program of High Education (grant nos. 20060001059 and 2000000127 to X.P.D.), and Peking University.





































