



Introduction
The trilobite subfamily Synphoriinae appeared in the Silurian but attained its greatest diversity in the Early to Middle Devonian. Silurian records, ranging in age from Sheinwoodian to Gorstian, are known from regions of the United States that were situated on or close to the southern margin of the Laurentian paleocontinent, as well as from Wales and northern England (Avalonia), Sweden (Baltica), and the Prague Basin of Czechia (the Bohemian paleocontinent or Perunica) (Holloway, Reference Holloway1981; Storey et al., Reference Storey, Thomas and Owens2016). In the Devonian, the subfamily is known from the upper Lochkovian to Eifelian, with its center of radiation in the eastern United States and southeastern Canada (southern Laurussia) (Lespérance and Bourque, Reference Lespérance and Bourque1971; Lespérance, Reference Lespérance1975; excluding Roncellia and Forillonaria of those authors, respectively, which belong to the Dalmanitinae, see Campbell, Reference Campbell1977). Other Devonian occurrences are in Bolivia (Holloway and Carvalho, Reference Holloway and Carvalho2010) and Colombia (Morzadec et al., Reference Morzadec, Mergl, Villarroel, Janvier and Racheboeuf2015), which were situated on the northwestern margin of Gondwana across the Rheic Ocean from Laurussia. We here augment the Devonian record with a new synphoriine, Needmorella simoni n. gen. n. sp., from the Needmore Shale of West Virginia. The new genus is not very similar to other Devonian Synphoriinae, with many of the differences from them reflecting less-derived character states that are present in Silurian members of the subfamily.
Stratigraphy, fauna, and age
The Needmore Shale of the central Appalachian Basin, named by Willard (Reference Willard, Willard, Swartz and Cleaves1939, p. 149), crops out in the Valley and Ridge physiographic province in central Pennsylvania, western Maryland, northeastern West Virginia, and western Virginia. It was previously referred to as the Onondaga Formation (e.g., Willard, Reference Willard1936), with which it is partly correlated in the New York sequence, and in Maryland, it was also called the Onondaga Member of the Romney Formation (Kindle in Prosser et al., Reference Prosser, Kindle and Swartz1913a, p. 48, 49). The Needmore overlies the Oriskany Sandstone unconformably or disconformably and is in turn overlain conformably by the Marcellus Shale, at the base of which is the Tioga Ash Bed. In Virginia and West Virginia, the Needmore is typically 20–30 m thick (Enomoto et al., Reference Enomoto, Coleman, Haynes, Whitmeyer, McDowell, Lewis, Spear and Swezey2012, p. 3) and consists of dark- to greenish-gray shales and siltstones that are commonly calcareous and contain calcareous nodules and discrete beds of argillaceous limestone (Hunt et al., Reference Hunt, McDowell, Blake, Toro and Dinterman2017, p. 62; Haynes et al., Reference Haynes, Pitts, Doctor, Diecchio and Blake2018, p. 14). The formation is considered to have been deposited in progressively deeper environments from ramp to basin margin, representing the beginning of foreland basin subsidence associated with the Acadian Orogeny (Haynes et al., Reference Haynes, Pitts, Doctor, Diecchio and Blake2018, p. 13, 43).
A diverse fossil fauna, including corals, bryozoans, brachiopods, bivalves, gastropods, cephalopods, tentaculitids, ostracods, and trilobites, was described by Kindle (Reference Kindle1912) and Prosser et al. (Reference Prosser, Kindle, Ulrich and Bassler1913b) from the ‘Onondaga’ (= Needmore Shale) in Pennsylvania, Maryland, Virginia, and West Virginia, with most of the named species having been previously documented from the New York sequence. Willard (Reference Willard, Willard, Swartz and Cleaves1939, table 22, p. 156–160, pls. 16, 17, 22, 24) and Butts (Reference Butts1940, p. 303–305; 1941, pls. 114–117) gave extensive lists of the Needmore fauna in Pennsylvania, Virginia, and West Virginia, and also illustrated some taxa; a similar list from the formation in West Virginia was given by Woodward (Reference Woodward1943). Cephalopods from the formation in Virginia and West Virginia were described by Miller (Reference Miller1938) and House (Reference House1978), as were ostracods from Pennsylvania and West Virginia by Swartz and Swain (Reference Swartz and Swain1941), rugose corals from Pennsylvania, Virginia, and West Virginia by Oliver (Reference Oliver1989), and the trilobite Viaphacops variabilis (Eldredge, Reference Eldredge1973) (= ‘Phacops cristata variabilis’) from Pennsylvania by Eldredge (Reference Eldredge1973). Other authors who discussed the Needmore fauna are de Witt and Colton (Reference de Witt and Colton1964), Inners (Reference Inners, Dennison, Hasson, Hoskins, Jolley and Sevon1979), and Newton (Reference Newton, Dennison, Hasson, Hoskins, Jolley and Sevon1979), the last recognizing the ‘Phacops,’ Planolites-Chondrites, and Orbiculoidea-Pacificocoelia biofacies, corresponding to increasing depths of deposition. Burns (Reference Burns1991) listed a diverse fauna from the formation at four localities in West Virginia and illustrated some of the taxa in drawings. The specimens of Needmorella simoni n. gen. n. sp. described herein were collected from one of his localities.
Viaphacops variabilis is the only trilobite previously named based on specimens from the Needmore Shale. Other trilobites from the formation were illustrated by Kindle (Reference Kindle1912), Butts (Reference Butts1941), and Burns (Reference Burns1991). Families represented are Proetidae, Aulacopleuridae, Brachymetopidae, Phacopidae, Dalmanitidae, Odontopleuridae, and Lichidae. Genera present, in current taxonomy and in addition to Viaphacops Maksimova, Reference Maksimova1972, include Australosutura Campbell & Goldring in Amos et al., Reference Amos, Campbell and Goldring1960, Eldredgeops Struve, Reference Struve1990, Coronura Hall and Clarke, Reference Hall and Clarke1888, Odontocephalus Conrad, Reference Conrad1840, Kettneraspis Prantl and Přibyl, Reference Prantl and Přibyl1949, and ‘Belenopyge’ Pek and Vaněk, Reference Pek and Vaněk1991. Some of these genera have been collected by the second author at the type locality of Needmorella simoni n. gen. n. sp. Confirming most species assignments of previous authors is beyond the scope of the present work.
The Needmore Shale was correlated by Ver Straeten (Reference Ver Straeten1996, p. 20, fig. 3; Reference Ver Straeten2007, fig. 18) with the Esopus, Schoharie, and Onondaga formations of New York. Conodonts from the Needmore were identified by Harris et al. (Reference Harris, Stamm, Weary, Repetski, Stamm and Parker1994) as indicating an interval from the Polygnathus costatus patulus Biozone to the lower Polygnathus costatus costatus Biozone. The goniatite Mithraxites buttsi (Miller, Reference Miller1938), documented by House (Reference House1978) as Foordites buttsi from high in the Needmore, was correlated by Becker and House (Reference Becker and House2000, p. 122) with the mid-Eifelian Pinacites jugleri Biozone. Monazite crystals from the Tioga ash bed B, at the base of the overlying Marcellus Shale, have U/Pb ages of 390.0 ± 0.5 Ma (Roden et al., Reference Roden, Parrish and Miller1990). All of this evidence indicates that the Needmore Shale ranges in age from late Emsian to mid-Eifelian.
Materials and methods
The specimens of Needmorella simoni n. gen. n. sp. were collected from the Needmore Shale in a cutting on the western side of Carpers Pike (State Route 259), ~ 770 m southwest of the intersection of Capon Springs Road and ~ 9 km northeast of Wardensville, Hampshire County, northeastern West Virginia (latitude 39.152758, longitude −78.541004). This is locality 18 of Burns (Reference Burns1991). The geological map of Dean et al. (Reference Dean, Kulander and Lessing1985) places the locality at the base of the combined Needmore and Marcellus shales as depicted, just above the top of the Oriskany Sandstone. The specimens are preserved as internal and external molds in greenish gray mudstone that weathers to buff, orange-gray, or pale gray in color. The specimens have been deformed tectonically to varying degrees by shearing and crushing.
Morphological terminology follows Whittington and Kelly (Reference Whittington, Kelly and Kaesler1997).
Repository and institutional abbreviation
Specimens are housed in the Department of Paleobiology, National Museum of Natural History (Smithsonian Institution), Washington, DC (USNM PAL).
Systematic paleontology
Family Dalmanitidae Vogdes, Reference Vogdes1890
Subfamily Synphoriinae Delo, Reference Delo1935
Remarks
Silurian members of the subfamily are Delops Rickards, Reference Rickards1965 (= Lygdozoon Holloway, Reference Holloway1981), Rickardsia Storey, Thomas, and Owens, Reference Storey, Thomas and Owens2016 (= Delops sensu Holloway, Reference Holloway1981 and Šnajdr, Reference Šnajdr1982), and Struveria Rickards, Reference Rickards1965. As type species of Delops, Rickards (Reference Rickards1965) designated ‘Phacops’ obtusicaudatus Salter, Reference Salter1849 from the upper Homerian of northern England, selecting as lectotype of the species an incomplete and strongly deformed internal mold of a cephalon (Rickards, Reference Rickards1965, pl. 84, fig. 1; Storey et al., Reference Storey, Thomas and Owens2016, fig. 5A), the only other former syntype, a pygidium, having been lost. The lectotype is an unsatisfactory basis for a species, showing few distinctive features, and Rickards (Reference Rickards1965) seems to have primarily based his concept and diagnosis of Delops not on Delops obtusicaudatus but on two other forms that he assigned to the genus, namely ‘Phacops (Dalmania)’ nobilis Thomas, Reference Thomas1900 and a newly erected subspecies ‘Delops’ nobilis marri Rickards, Reference Rickards1965. Additional and better preserved specimens of Delops obtusicaudatus, from the same region as the type locality and from Wales, were documented by Storey et al. (Reference Storey, Thomas and Owens2016, fig. 5) who considered that they indicate that Delops is a senior subjective synonym of Lygdozoon, and that the species Delops nobilis and Delops marri (the latter regarded by those authors as a junior synonym of ‘Dalmanites’ mobergi Hede, Reference Hede1915 from the Homerian of Sweden) belong to a separate genus which they named Rickardsia.
We exclude from Silurian representatives of the Synphoriinae the species ‘Phacops’ weaveri Salter, Reference Salter1849, from the Telychian of Gloucestershire, England. This species was assigned to Lygdozoon by Curtis and Lane (Reference Curtis and Lane1998, p. 69, pl. 8, figs. 1, 10–12) and to ‘Delops sensu lato’ by Storey et al. (Reference Storey, Thomas and Owens2016, p. 4, 5). However, its pattern of pygidial segmentation is typical of members of the Dalmanitinae, in particular the pleural furrows with abrupt anterior slopes and longer, more gentle posterior slopes, and it belongs to that subfamily (see Holloway, Reference Holloway1981, p. 697, 698). Also belonging to the Dalmanitinae, as indicated particularly by the spacing and form of the glabellar apodemes, is the incomplete cranidium described by Waisfeld et al. (Reference Waisfeld, Toro and Brussa1988, p. 316, pl. 2, fig. 14) as ‘Synphoriinae gen. et sp. indet.,’ from late Silurian strata of the Los Espejos Formation in the Precordillera of Argentina. The cranidium is probably conspecific with one or more of the other specimens assigned to Dalmanites Barrande, Reference Barrande1852 by those authors from the same stratigraphic section.
Of the Devonian members of the Synphoriinae, we here discuss only Synphoria Clarke, Reference Clarke, Ulrich, Clarke, Scofield and Winchell1894, Anchiopsis Delo, Reference Delo1935, and Synphoroides Delo, Reference Delo1940, other genera being very different from Needmorella n. gen. Synphoria, with type species Synphoria stemmata (Clarke, Reference Clarke1900) from the Glenerie Formation of New York, occurs in the Pragian and possibly Emsian in eastern North America. The type species was also reported by Morzadec et al. (2015, fig. 9A–C) from the upper Emsian of Colombia, based on several pygidia, but the specimens belong to the Dalmanitinae rather than to the Synphoriinae. Authorship of Synphoria has generally been considered to date from Clarke (Reference Clarke1900) (e.g., Reed, Reference Reed1927, p. 343; Delo, Reference Delo1935, p. 412; 1940, p. 65; Hupé, Reference Hupé and Piveteau1953, p. 235; Richter et al., Reference Richter, Richter, Struve and Moore1959, p. 472; Lespérance and Bourque, Reference Lespérance and Bourque1971, p. 194; Lespérance, Reference Lespérance1975, p. 103; Jell and Adrain, Reference Jell and Adrain2003, p. 450; Morzadec et al., Reference Morzadec, Mergl, Villarroel, Janvier and Racheboeuf2015, p. 346). However, the name was first used by Clarke (Reference Clarke, Ulrich, Clarke, Scofield and Winchell1894) (to whom authorship was attributed by Reed, Reference Reed1905, p. 173, and Rennie, Reference Rennie1930, p. 330) as a subdivision of Dalmanites to incorporate several taxa (Odontocephalus, Corycephalus Hall and Clarke, Reference Hall and Clarke1888, and Coronura) that at that time were also regarded as subgenera of Dalmanites, and that Clarke considered to be characterized by coalescence of L3 and L2 (‘first and second lateral glabellar lobes,’ respectively, in his terminology). This earlier usage of Synphoria by Clarke was considered to be nomenclatorially invalid by Delo (Reference Delo1940) and Richter et al. (Reference Richter, Richter, Struve and Moore1959) because they considered it to have been applied to a collective group of uncertain taxonomic level; for the same reason, Richter and Richter (Reference Richter and Richter1942, p. 175) considered that the name Synphoria was not valid at all. However, Clarke (Reference Clarke, Ulrich, Clarke, Scofield and Winchell1894, p. 733) clearly stated that “… for such a subgeneric division a designation is needed (e.g., Synphoria).” Even in the absence of that statement by Clarke (Reference Clarke, Ulrich, Clarke, Scofield and Winchell1894), the name must be regarded as having been applied to a subgenus because it was proposed for a genus-group division of the genus Dalmanites (ICZN, 1999, Article 10.4). No named species were assigned to Synphoria by Clarke (Reference Clarke, Ulrich, Clarke, Scofield and Winchell1894) but that does not affect the availability of a generic or subgeneric name established before 1931 (ICZN, 1999, Article 67.2.2).
In the case of Anchiopsis, there are difficulties in its interpretation and differentiation from Synphoria. When proposing Anchiopsis, with type species ‘Calymene’ anchiops Green, Reference Green1832 from the Schoharie Formation (late Emsian) of New York, Delo (Reference Delo1935) did not compare it with Synphoria. He later stated (Delo, Reference Delo1940, p. 83) that Anchiopsis “… marks the climax of [glabellar] lobal coalescence and elevation in the Synphoriinae,” but he also noted that Synphoria shows similar coalescence of the lobes. He differentiated Synphoria from Anchiopsis by the less-convex cephalon, less-swollen L2 and L3, and deeper S1 and S2 of the former; however, we can see no consistent differences between the two genera in these features. Anchiopsis anchiops is generally considered to possess a robust occipital spine, short genal spines, and a long pygidial mucro (e.g., Hall and Clarke, Reference Hall and Clarke1888, pl. 9, figs. 1–5, 12, pl. 10, figs. 3–10, 12, 13; Ludvigsen, Reference Ludvigsen1979, fig. 51A, B; Whiteley et al., Reference Whiteley, Kloc and Brett2002, fig. 5.2B, pl. 120), and these characters were regarded as diagnostic of the genus by Richter et al. (Reference Richter, Richter, Struve and Moore1959, p. 469, 470; = ‘Anchiopella’ Reed, Reference Reed1907 of their usage) and Lespérance and Bourque (Reference Lespérance and Bourque1971, p. 202), although Anchiopsis armata (Hall, Reference Hall1861) has elongated genal angles lacking spines (Hall and Clarke, Reference Hall and Clarke1888, pl. 9, figs. 7–9; Morzadec et al., Reference Morzadec, Mergl, Villarroel, Janvier and Racheboeuf2015, fig. 8A–D). Genal spines and a mucro can also be present in Synphoria; e.g., Synphoria dolbeli (Clarke, Reference Clarke1907), see Clarke (Reference Clarke1908, pl. 8, figs. 1–7), and Synphoria sopita Lespérance, Reference Lespérance1975, these species probably synonyms in our opinion. In the holotype of Anchiopsis anchiops, an articulated dorsal exoskeleton illustrated by Hall (Reference Hall1876, pl. 9, fig. 12 of Crustacea) and Hall and Clarke (Reference Hall and Clarke1888, pl. 9, fig. 13), the occipital ring and pygidium are broken posteriorly, so it cannot be verified whether an occipital spine and mucro were originally present, as thought by Hall and Clarke (Reference Hall and Clarke1888, explanation to pl. 9), although what is preserved of the pygidial margin suggests that it is unlikely that a mucro was present. The genal angles of the cephalon are rounded rather than spinose in the holotype. Apparently very similar to this specimen are the cephalon, lacking occipital and genal spines, and the pygidium, lacking a mucro, as illustrated by Hall (Reference Hall1876, pl. 9, fig. 11, pl. 10, fig. 12 of Crustacea) and Hall and Clarke (Reference Hall and Clarke1888, pl. 9, fig. 11, pl. 10, fig. 2), both specimens also from the Schoharie Formation. These specimens are, respectively, the holotype of ‘Dalmanites (Chasmops)’ anchiops var. sobrinus Hall and Clarke, Reference Hall and Clarke1888 and a syntype of ‘Asaphus’ laticostatus Green, Reference Green1832, the latter being regarded by Hall (Reference Hall1876) and Hall and Clarke (Reference Hall and Clarke1888) as a synonym of Anchiopsis anchiops. Lespérance and Bourque (Reference Lespérance and Bourque1971) considered ‘Dalmanites (Chasmops)’ anchiops var. sobrinus to be a possible synonym of their new subspecies Synphoria stemmata compacta Lespérance and Bourque, Reference Lespérance and Bourque1971, which is from the stratigraphically older (Pragian) Glenerie Formation and based on paralectotypes of the nominotypical subspecies of Synphoria stemmata, of which it is probably a synonym. We consider that ‘Dalmanites (Chasmops)’ anchiops var. sobrinus as well as ‘Asaphus’ laticostatus might be synonyms of Anchiopsis anchiops, that most specimens that have been assigned to Anchiopsis are not conspecific with the holotype of Anchiopsis anchiops, and that Anchiopsis might be a junior synonym of Synphoria. These issues can only be resolved by thorough revision of Anchiopsis including restudy of the type specimens of the type species as well as those of ‘Dalmanites (Chasmops)’ anchiops var. sobrinus and ‘Asaphus’ laticostus, but that is beyond the scope of the present work.
Lespérance and Bourque (Reference Lespérance and Bourque1971, pl. 27, figs. 1–3) assigned to Anchiopsis anchiops an exfoliated cephalon lacking a genal spine and a pygidium, from the ‘upper Grande Grève Formation’ (= Shiphead or Indian Cove formations in current terminology, see Lespérance, Reference Lespérance1980; Pragian or early Emsian) of Quebec. We consider that these specimens belong to Synphoria dolbeli, of which the cephalon is a syntype (see Kilfoyle, Reference Kilfoyle1954, p. 527) despite arguments to the contrary by Lespérance and Bourque (Reference Lespérance and Bourque1971, p. 204) who referred to it incorrectly as a ‘hypotype’ of that species. Apparent differences between that cephalon and the other from the same formation illustrated by those authors (Lespérance and Bourque, Reference Lespérance and Bourque1971, pl. 27, figs. 6, 7) as S. dolbeli, in the shape of the frontal lobe and the length of the genal field in front of the eye, can be accounted for by tectonic flattening of the first specimen, and the more forwardly tilted orientation of the second (compare with the stereo-pair photographs of the second cephalon by Lespérance, Reference Lespérance1975, pl. 1, fig. 1). The pygidium figured as Anchiopsis anchiops by Lespérance and Bourque (Reference Lespérance and Bourque1971) resembles in all observable characters the other pygidia figured by them as Synphoria dolbeli (Lespérance and Bourque, Reference Lespérance and Bourque1971, pl. 27, figs. 8, 9). The small pygidium figured as ‘Anchiopsis aff. A. anchiops’ by Lespérance and Bourque (Reference Lespérance and Bourque1971, pl. 27, fig. 4) does not belong to the Synphoriinae but to the Dalmanitinae.
Synphoroides, with type species Synphoroides biardi (Clarke, Reference Clarke1907), from the upper Rensselaeria zone of the ‘Grande Grève Formation’ (Lespérance and Bourque, Reference Lespérance and Bourque1971, text-fig. 2), now Shiphead Formation (Pragian) of Quebec, was considered by Delo (Reference Delo1940, p. 78, 79) and Lespérance and Bourque (Reference Lespérance and Bourque1971, p. 193) to be distinguished especially by a bifid or trifid anterior cephalic process. The lectotype cephalon of Synphoroides biardi, illustrated by Clarke (Reference Clarke1908, pl. 6, fig. 8; reproduced by Delo, Reference Delo1940, pl. 9, fig. 5, and Linsley, Reference Linsley1994, pl. 308, fig. 2), has a bifid anterior process, together with several successively smaller marginal crenulations laterally. In the other cephala and cephalic fragments figured as Synphoroides biardi by Clarke (Reference Clarke1908), the anterior process is longer and trifid. Clarke (Reference Clarke1908, p. 264) and Delo (Reference Delo1940, p. 128) attributed this difference to breakage of the medial branch of the process in the lectotype but, judging from the illustrations, this appears very unlikely. The lectotype also differs from the other complete cephalon illustrated by Clarke (Reference Clarke1908, pl. 6, fig. 12; also Delo, Reference Delo1940, pl. 9, fig. 4, and Linsley, Reference Linsley1994, pl. 308, fig. 6) in that the glabella is narrower (tr.) across the occipital ring; the eye is larger, reaching almost to the posterior border furrow and extending closer to the lateral border furrow anterolaterally; and the genal spine is longer. Hence, if Clarke's (Reference Clarke1908) illustrations are accurate, it appears that the type specimens could include more than one species. Delo (Reference Delo1940) assigned five species to Synphoroides apart from the type, but two of them (‘Dalmanites’ griffoni Clarke, Reference Clarke1907 and ‘Asaphus’ pleuroptyx Green, Reference Green1832; the latter well illustrated by Whiteley et al., Reference Whiteley, Kloc and Brett2002, pl. 90) were rejected by Lespérance and Bourque (Reference Lespérance and Bourque1971, p. 194), and we agree that these belong to the Dalmanitinae rather than to the Synphoriinae. Also belonging to the Dalmanitinae are ‘Dalmanites (Synphoria)’ esnoufi Clarke, Reference Clarke1907 and ‘Dalmanites’ multiannulatus Ohern in Bassler et al., Reference Bassler, Maynard, Ohern, Schuchert, Swartz and Ulrich1913. We tentatively retain in Synphoroides two other species, ‘Dalmanites’ dolphi Clarke, Reference Clarke1893 (see Delo, Reference Delo1940, pl. 9, fig. 10), from the Port Jervis Formation (Pragian) of New York, and Synphoroides? vetustus Lespérance, Reference Lespérance1975 from the Roncelles Formation (Lochkovian?) of Quebec. However, the anterior cephalic border is poorly preserved in each and they might belong to Synphoria. The identification by Haas (Reference Haas1969, p. 651, pl. 83, figs. 5–8) of Synphoroides biardi from the upper Pragian of central Chihuahua, northern Mexico, was based on fragmentary cephala and a pygidium that, judging especially from the pygidium, belong to the Dalmanitinae rather than to the Synphoriinae.
Genus Needmorella new genus
Type species
Needmorella simoni n. gen. n. sp.
Diagnosis
The diagnosis given for the species serves as diagnosis for the genus due to monotypy.
Etymology
After the Needmore Shale in which the genus occurs, together with the Latin diminutive suffix –ella; gender feminine.
LSID
urn:lsid:zoobank.org:act:8ECB7853-942C-4D25-8364-A11872CFEDD7.
Remarks
Needmorella n. gen. is not very similar to other Devonian synphoriines. Synphoria, Synphoroides, and Anchiopsis (as the latter is conventionally interpreted; see preceding remarks on the subfamily) differ in that the anterior cephalic border is longer (sag., exsag.) and bears multiple crenulations or spines; the glabella is broader across L1 and does not expand as strongly forward; S2 contains an apodemal pit adaxially and is shallow or absent abaxially, so that L2 and L3 tend to coalesce; S2 and S3 extend closer to the sagittal line, the central area of the glabella being narrower; L2 and especially L3 are more inflated, at least in Synphoria; genal spines are much shorter or absent; the posterior pleural bands are not more inflated and expanded abaxially than the anterior bands in the thorax and pygidium; and the interpleural furrows are very weak on the pygidium. In addition, a marked subocular groove and prominent subocular ridge are present in at least Synphoria and Synphoroides (see Clarke, Reference Clarke1908, pl. 6, fig. 8; Lespérance, Reference Lespérance1975, pl. 1, figs. 8, 10, pl. 2, figs. 6, 7); and on the cephalic doublure of Synphoria, the vincular furrow lies in front of instead of behind the prominent rounded ridge running subparallel to the cephalic margin (Clarke, Reference Clarke1900, pl. 2, figs. 1, 2; Lespérance and Bourque, Reference Lespérance and Bourque1971, pl. 26, fig. 4, pl. 27, fig. 6), this difference probably a result of the longer anterior border in Synphoria. Even more dissimilar to Needmorella n. gen. are Coronura and Odontocephalus, which occur together with it in the Needmore Shale.
In the very short (sag., exsag.), unmodified anterior cephalic border, S2 that is well defined abaxially, and the well-impressed pygidial interpleural furrows, Needmorella n. gen. resembles the Silurian synphoriines Delops, Rickardsia, and Struveria rather than other Devonian members of the subfamily. These similarities reflect less-derived character states. There are similarities with Rickardsia also in the pygidium with inflated posterior pleural bands that project slightly distally, although this feature is not as strongly developed in that genus (Rickards, Reference Rickards1965, pl. 84, figs. 6, 7; Storey et al., Reference Storey, Thomas and Owens2016, fig. 7O–R). The similarities in all of these characters with Silurian members of the subfamily suggest that Needmorella n. gen. was not descended from the most recent common ancestor of Synphoria and its close allies but belongs to a morphologically more conservative lineage that did not possess the derived character states, and that persisted from the Silurian into the Devonian.
Needmorella simoni new genus new species
Figures 1–3
- Reference Kindle1912
Cryphaeus cf. boothi var. calliteles (Kayser) (sic); Kindle, p. 108, pl. 10, figs. 14, 15, pl. 11, fig. 1.
- Reference Kindle1912
Phaethonides gemmaeus Hall and Clarke; Kindle, p. 111, pl. 13, fig. 12; in part, not pl. 13, fig. 11 = Australosutura gemmaea?
- ?Reference Willard, Willard, Swartz and Cleaves1939
Greenops boothi, var. calliteles; Willard, p. 159 (list).
- Reference Butts1941
Dalmanites aspectans Conrad? (sic); Butts, pl. 116, figs. 11, 12.
- ?Reference Woodward1943
Greenops boothi var. calliteles (Green); Woodward, p. 302 (list).
- Reference Burns1991
Dalmanites sp.; Burns, fig. 15 on p. 85, fig. 4 on p. 89.
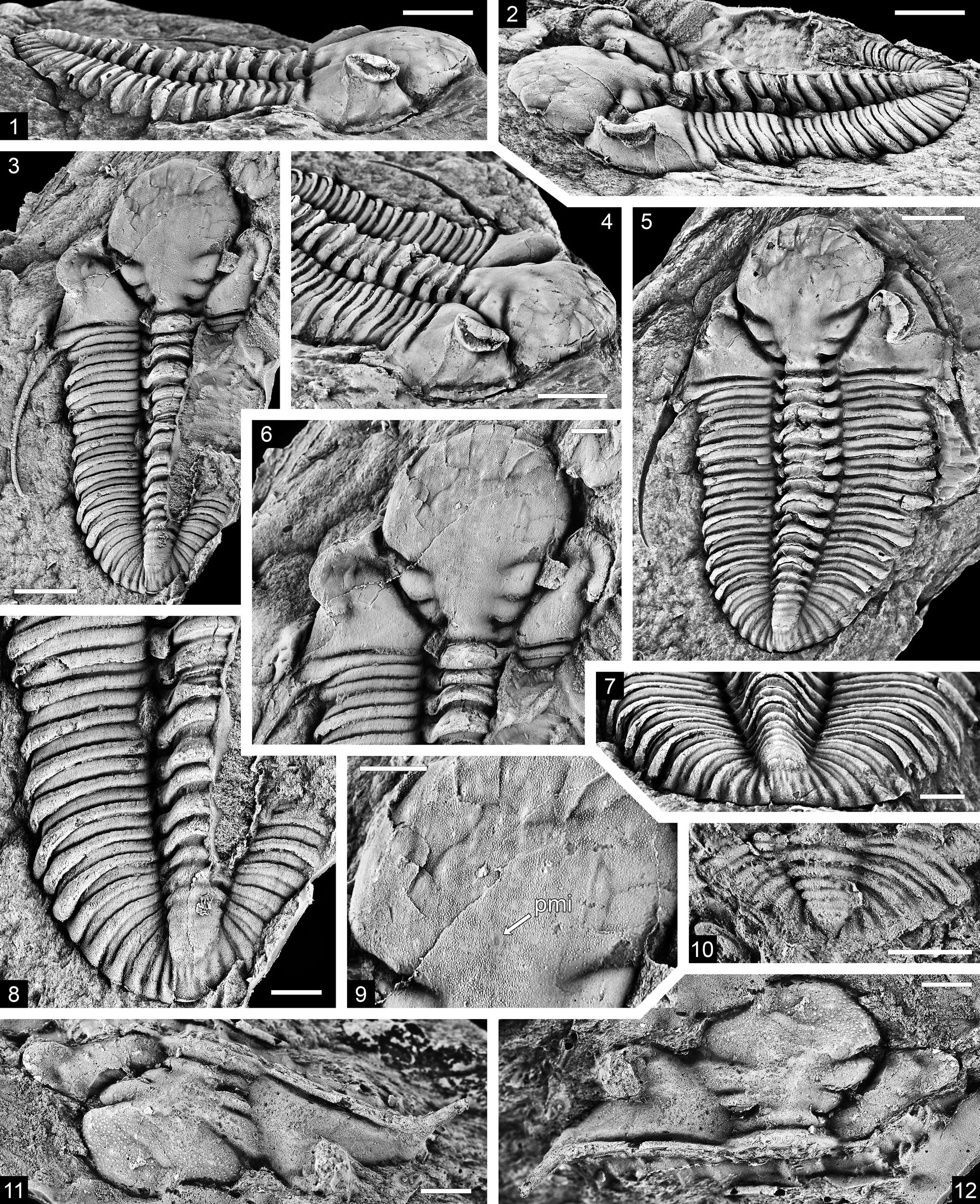
Figure 1. Needmorella simoni n. gen. n. sp., Needmore Shale (Emsian–Eifelian), northeastern West Virginia: (1–9) holotype, counterpart molds of dorsal exoskeleton, USNM PAL 777869A, B: (1, 4, 5, 7) internal mold, USNM PAL 777869B, lateral view, oblique view of cephalon and thorax, dorsal view, and posterodorsal view of pygidium and posterior thorax; (2, 3, 6, 8, 9) external mold, USNM PAL 777869A, latex cast, dorsolateral and dorsal views, dorsal view of cephalon and anterior thorax, dorsal view of thorax and pygidium, and detail of glabella in dorsal view showing finely granulose sculpture and posterior median impression (pmi) on frontal lobe; (10) small pygidium, USNM PAL 777876, latex cast of external mold, dorsal view; (11, 12) cranidium, USNM PAL 777873, latex cast of external mold, oblique and dorsal views. Scale bars = 5 mm (1–5), 2 mm (6–12).

Figure 2. Needmorella simoni n. gen. n. sp., Needmore Shale (Emsian–Eifelian), northeastern West Virginia: (1) cranidium, USNM PAL 777877A, latex cast of external mold, dorsal view; (2) cranidium, USNM PAL 777879, internal mold, dorsal view; (3) small cranidium, USNM PAL 777883, internal mold, dorsal view; (4) cephalon, USNM PAL 777872, latex cast of external mold, detail of doublure, ventral view; (5) thoracopygon, USNM PAL 777889B, internal mold, dorsal view; (6) small cranidium, USNM PAL 777884A, latex cast of external mold, dorsal view; (7) librigenal doublure, USNM PAL 777888, latex cast of external mold, ventral view; (8) small cranidium, USNM PAL 777882B, internal mold, dorsal view; (9, 10, 13) cranidium, USNM PAL 777875, latex cast of external mold, dorsal, oblique and anterodorsal views; (11, 14, 17) pygidium, USNM PAL 777890A, latex cast of external mold, lateral, posterodorsal and dorsal views; (12, 15) counterpart molds of small cranidium, USNM PAL 777885A, B, dorsal view: (12) internal mold, USNM PAL 777885B; (15) external mold, USNM PAL 777885A, latex cast; (16) cephalon, USNM PAL 777871, internal mold, oblique view. Scale bars = 2 mm.

Figure 3. Needmorella simoni n. gen. n. sp., Needmore Shale (Emsian–Eifelian), northeastern West Virginia: (1, 4) counterpart molds of cranidium, USNM PAL 777880A, B, dorsal view: (1) internal mold, USNM PAL 777880B; (4) external mold, USNM PAL 777880A, latex cast; (2, 3) counterpart molds of cranidium, USNM PAL 777874A, B, dorsal view: (2) external mold, USNM PAL 777874A, latex cast; (3) internal mold, USNM PAL 777874B; (5) cranidium, USNM PAL 777881, internal mold, dorsal view; (6) pygidium, USNM PAL 777891, latex cast of external mold, dorsal view; (7) small pygidium, USNM PAL 777893A, latex cast of external mold, dorsal view; (8) cranidium, USNM PAL 777878B, internal mold, dorsal view; (9) cephalon, USNM PAL 777870, internal mold, oblique view; (10) small cranidium, USNM PAL 777886B, internal mold, dorsal view; (11) pygidium, USNM PAL 777887A, latex cast of external mold, posterior view; (12, 13) pygidium, USNM PAL 777892, latex cast of external mold, posterior and dorsal views. Scale bars = 2 mm.
Type specimens
Holotype, counterpart external and internal molds of articulated dorsal exoskeleton, USNM PAL 777869A, B (Fig. 1.1–1.9). Paratypes: cephala, USNM PAL 777870–777872; cranidia, USNM PAL 777873–777875, 777877–777886; librigenal doublure, USNM PAL 777888; thoracopygon, USNM PAL 777889; pygidia, USNM PAL 777876, 777887–777894.
Diagnosis
Anterior cephalic border very short (sag., exsag.), not modified by spines or crenulations; genal spines very long, slender, and curved; glabella expanding strongly forward in front of L1, approximately twice as wide across posterior part of frontal lobe as across L1; S2 lacking apodemal pit adaxially, meeting axial furrow abaxially; S2 and S3 extending adaxially less than half way to sagittal line, so that central area of glabella is very broad; eye of moderate size, posterior edge situated well forward opposite front of L2; cephalic doublure with vincular furrow situated behind a rounded (sag., exsag.) marginal ridge anteriorly and laterally. Pygidium nonmucronate; interpleural furrows almost as deep as pleural ones; posterior pleural bands on pygidium and posterior half of thorax increasing in convexity abaxially and expanding at expense of anterior bands, projecting slightly distally.
Occurrence
Needmore Shale, Hampshire County, northeastern West Virginia.
Description
Cephalon broadly parabolic in outline, its anterior margin lacking crenulations or spines (Fig. 2.4, 2.7); cephalic proportions difficult to determine due to deformation, but specimens that appear least deformed suggest that width across base of genal spines is ~170–200% length (sag.). Glabella subpentagonal in outline but well-rounded anteromedially, comprising ~25% cephalic width posteriorly (excluding genal spines), slightly narrower across L1 than across occipital ring, expanding almost uniformly from S1 to posterior part of frontal lobe except for slight outward bulge across anterior half of L3, maximum widths across L1, L2 and L3 approximately in ratio 1:1.2:1.8. Medial portion of occipital ring (approximately half total width) gently rounded (sag., exsag.) and of uniform length, lateral portion shorter and deflected forward abaxial to occipital apodemal pit; in transverse profile, occipital ring not evenly convex but strongly arched medially and gently concave on flanks (Fig. 2.13); occipital furrow transverse medially, deflected backward toward apodemal pit beyond which it is directed anterolaterally. L1 similar in appearance to occipital ring but a little shorter (sag., exsag.), less rounded (sag., exsag.) in median portion and with weak, slightly depressed lateral nodes; deep lateral portion of S1 curving forward adaxially. L2 ~150% as long (exsag.) as L1; S2 transverse or directed weakly forward adaxially, not extending as close to sagittal line as S1. L3 almost twice as long (exsag.) abaxially than adaxially where it is a little longer than L2; S3 directed obliquely backward at ~60–70° to sagittal line, its inner end more or less in line (exsag.) with that of S2. Frontal lobe comprising half sagittal glabellar length or more, subrhombic to subelliptical in outline, ~140–160% as wide as long (sag.), with small posterior median impression situated slightly behind its maximum width (Fig. 1.9). Axial furrow shallow adjacent to occipital ring and L1, well impressed from S1 to widest part of frontal lobe. Palpebral lobe convex (tr.), semicircular in lateral outline, situated approximately half its length from posterior border furrow, farther from sagittal line posteriorly than anteriorly; palpebral furrow with strongest curvature at midlength; visual surface poorly preserved due to crushing, lens arrangement indeterminate. Posterior branch of facial suture crossing gena in broad, convex-forward arc lateral to eye, meeting lateral border furrow more or less opposite ɛ (Figs. 1.5, 2.16, 3.9); anterior branch running parallel to axial furrow before curving sharply adaxially across border furrow opposite lateral extremity of frontal lobe. Fixigenal field gently convex (exsag., tr.) adaxial to palpebral lobe and posteromedially where it is elevated above posterior border, becoming gently concave (exsag.) posterolaterally; librigenal field steeply sloping, much wider than lateral border including at front of eye (Fig. 1.1, 1.4). Posterior border approximately half as long (exsag.) proximally as lateral extremity of occipital ring, expanding slightly abaxially as far as fulcrum, thereafter expanding more strongly to approximately twice its proximal length, which it maintains for a short distance to base of genal spine (Fig. 1.6); posterior border furrow curving backward slightly beyond fulcrum, dying out distally before reaching lateral border furrow. Lateral border narrow, of uniform width, its dorsal surface gently convex; lateral border furrow shallow (Figs. 2.16, 3.9), obscured by compression fracture in holotype (Fig. 1.1, 1.4, 1.5). Genal spine initially diverging at ~30° to sagittal line, gradually tapering and curving backward and adaxially, its tip lying more or less opposite tenth thoracic segment. Cephala and cranidia of small and moderate size with sculpture of rather sparse, coarse granules with interspersed scattered tubercles, the latter mainly on glabella and palpebral lobe; in cranidia of moderate size, tubercles mainly confined to frontal lobe but in small cranidia are also sparsely developed on L2 and L3, with apparent pairing of some tubercles here and on frontal lobe (Figs. 2.3, 2.6, 2.8, 3.4, 3.5). Sculpture on largest cephalon and cranidium consisting of very dense, fine granules, possibly superimposed on weak remnants of tubercles on frontal lobe (Figs. 1.9, 3.2).
Median part of cephalic doublure divided into rounded (sag., exsag) outer ridge and longer (sag., exsag.), weakly convex inner portion by broad, rather shallow vincular furrow running subparallel to outer cephalic margin (Fig. 2.7); inner portion of doublure narrowing strongly laterally and vincular furrow fading; posterolaterally, only very narrow, outer portion of doublure visible in ventral view, inner portion apparently here being strongly turned upward. Hypostomal suture broadly and evenly convex forward. Sculpture of dense, coarse granules on convex outer portion of doublure, and finer, sparse granules on posterior portion and in vincular furrow (Fig. 2.4, 2.7).
Thorax composed of 11 segments, of more or less uniform width across segments 1–6 and thereafter progressively narrowing, ~70% as wide posteriorly as anteriorly. Axis subparallel-sided on segments 1–5 or 6, thereafter progressively narrowing, ~60% as wide on last segment as on first, comprising ~30% total width of first segment; axial rings similar in form to occipital ring; axial furrow bowed outward on each segment. Pleural region strongly downturned at fulcrum, situated at ~66% distance from axial furrow; pleural furrows moderately deep; anterior and posterior pleural bands weakly rounded adaxial to fulcrum, approximately equal in length (exsag.) except very close to axial furrow where posterior bands are longer as pleural furrows curve forward; abaxial to fulcrum, posterior bands increase in length and greatly in convexity.
Pygidium 160% to a little more than 200% as wide as long; maximum width level with third or fourth axial ring; posterior margin more strongly curved medially than laterally. Axis strongly convex, comprising ~25% maximum pygidial width anteriorly and 75–85% sagittal length, narrowing uniformly backward to well-defined, broadly rounded terminus; seven axial rings plus short (sag.) terminal piece; first ring very prominent, strongly arched medially and concave on flanks like thoracic rings; subsequent rings successively less prominent and more evenly arched, second ring with weakly defined pseudoarticulating half ring; articulating furrow deep and expanded (sag., exsag.) medially, inter-ring furrows successively shorter (sag., exsag.) and shallower, weak behind third except that last appears as more distinct, paired impressions in at least some specimens (Fig. 1.5, 1.7, 1.10); articulating furrow and inter-ring furrows 1–3 or 4 containing apodemal pits laterally. Pleural region moderately convex (tr.) except close to margin where it is weakly concave; six (?), seven, or eight segments defined by inflated posterior pleural bands which, except for last few, form slight projections distally (Figs. 2.14, 3.11); shorter (exsag.) anterior pleural bands progressively reduced in development on successive segments, disappearing after fourth or fifth segment, beyond which posterior pleural bands are separated by combined pleural and interpleural furrows; holotype with weak, very narrow median postaxial ridge resembling adjacent posterior pleural bands (Fig. 1.7, 1.8); pleural and combined furrows deeper and wider (exsag.) than interpleural furrows except distally where interpleural ones deepen and expand slightly; interpleural and combined furrows extending more or less to pygidial margin, pleural furrows terminating just inside margin. Furrow behind articulating flange on anterior pleural margin extending abaxially almost to outer end of articulating facet (Fig. 2.11, 2.14, 2.17). Outer portion of doublure beneath distal projections of posterior pleural bands sloping downward and inward (Fig. 3.11). Sculpture of dense, fine granules on dorsal surface of pygidium, becoming coarser on distal ends of posterior pleural bands and outer portion of doublure.
Etymology
After the son of the second author.
LSID
urn:lsid:zoobank.org:act:28124EDC-79A6-41FE-900B-BC5DADA514EF.
Remarks
Cephala and cranidia range in length from 4.3–13.5 mm, and pygidia are ~6–11.5 mm wide. The only apparent morphological variation that can be attributed to ontogenetic change is in cephalic sculpture, rather sparse granules and scattered tubercles in small and medium-sized specimens being replaced in the largest specimens with finer, dense granules, possibly with weak remnants of tubercles on the frontal lobe. Variation in the width/length proportions of the pygidium appears to be unrelated to specimen size and is probably due largely to deformation.
We assign to Needmorella simoni n. gen. n. sp. the two cranidia and the dorsal exoskeleton with incomplete thorax and displaced pygidium from the ‘Onondaga’ (i.e., Needmore Shale) of West Virginia and Pennsylvania, illustrated by Kindle (Reference Kindle1912, pl. 10, figs. 14, 15, pl. 11, fig. 1) as ‘Cryphaeus cf. C. boothi var. calliteles’ = Bellacartwrightia calliteles (Green, Reference Green1837) (see Lieberman and Kloc, Reference Lieberman and Kloc1997); Kindle incorrectly attributed authorship of this species to Kayser (Reference Kayser1878) instead of to Green (Reference Green1837). Kindle mistakenly thought that the pygidium associated with the dorsal exoskeleton might belong to another species, suggesting that he had recognized the differences from the pygidium of Bellacartwrightia calliteles. Coming from the same region of Pennsylvania as the dorsal exoskeleton, and also belonging to N. simoni n. gen. n. sp., is the small pygidium assigned with question by Kindle (Reference Kindle1912, pl. 13, fig. 12) to ‘Phaethonides’ gemmaeus Hall and Clarke, Reference Hall and Clarke1888. The other pygidium illustrated by Kindle (Reference Kindle1912, pl. 13, fig. 11) as the latter species, from Virginia, differs markedly in proportions, pattern of segmentation, and tuberculate sculpture, and is probably correctly assigned to Australosutura gemmaea (Hall and Clarke, Reference Hall and Clarke1888). Inclusion of ‘Greenops boothi var. calliteles’ in the faunal lists of Willard (Reference Willard, Willard, Swartz and Cleaves1939) and Woodward (Reference Woodward1943) was probably based on the misidentification of this taxon by Kindle (Reference Kindle1912). Also belonging to N. simoni n. gen. n. sp. are the incomplete cephalon and pygidium, from the ‘Onondaga’ in Virginia, assigned by Butts (Reference Butts1941, pl. 116, figs. 11, 12) to ‘Dalmanites aspectans Conrad?’ (= Asaphus aspectans Conrad, Reference Conrad1841), type species of Coronura, and probably the dorsal exoskeleton and cephalon from West Virginia illustrated as Dalmanites sp. in the drawings of Burns (Reference Burns1991, fig. 15 on p. 85, fig. 4 on p. 89).
Acknowledgments
We thank E. Rhenberg (West Virginia Geological and Economic Survey) for kindly providing copies of some publications and for advice, and G.D. Edgecombe (Natural History Museum, London) and B.S. Lieberman (University of Kansas) for helpful reviews that improved the manuscript.
Declaration of competing interests
The authors declare none.



