


INTRODUCTION
Understanding the causes and consequences of co-infection remains one of the major challenges in disease biology. Although co-infection is the rule rather than the exception [Reference Brooker1], the consequences of multiple infections have been traditionally ignored not only by animal and human health scientists but also by wildlife ecologists. Nevertheless, there is an increasing interest to move from the ‘one-disease-one-parasite’ perspective to a more holistic view of hosts as ecosystems of parasites [Reference Pérez2], partially motivated by the health impact of co-occurring infections [Reference Pullan and Brooker3]. In fact, in such complex ‘host–parasite ecosystems’ a variety of both direct and indirect interactions between parasites and their hosts [Reference Pedersen and Fenton4, Reference Hatcher and Dunn5] must be taken into account. For example, acquired immunity to one pathogen species may have negative effects on a second species, but also can produce immunosuppression increasing infection susceptibility [Reference Telfer6]. On the other hand, direct competition for resources are also common [Reference Lello7]. Thus, both the host's immune response (top-down regulations [Reference Pedersen and Fenton4]) and competition between parasites (bottom-up regulations) have to be considered in order to explain the observed variability in susceptibility to a new infection [Reference Telfer8], infectiousness [Reference Telfer6] and even pathogen shedding [Reference Lass9] in an infected host.
One of the most relevant consequences of co-infection is its effect on disease severity. The cost of infection relies on both the number of species involved in the infection (see [Reference Miller10] for the combined effects of two pathogens, but [Reference Petney and Andrews11] for a broad community), and on pathogen burden [Reference Marinho12]. However, certain combinations could have a protective effect [Reference Nkuo-Akenji13], making it extremely difficult to assess the cost of infection by considering only a part of the whole host–pathogen community [Reference Boel14]. Nonetheless, most published information on the consequences of co-infection in wildlife [Reference Ezenwa and Jolles15] and humans [Reference Griffiths16] have been largely based on the combined effects of only two species, and thus whether the outcomes of such studies are conditioned by the number and type of pathogen species considered is yet to be determined.
In this work, we provide and example on the consequences of considering only a part of the complete pathogen community when assessing the cost of infection in adult feral cats (Felis silvestris catus). Cats are particularly suitable for our purposes since they do not store fat reserves in anticipation of food shortage (i.e. income breeders). An evaluation of the nutrient requirements of domestic cats has led to the conclusion that cats are metabolically attuned to a carnivorous diet [Reference McDonald, Rogers and Morris17] and hence a high protein diet limits the likelihood of obesity and deviations from a balanced body condition often results in disease [Reference Becvarova18]. Therefore, poor body condition in cats has often been linked to infections caused by viruses [Reference Gilot-Fromont19], helminths [Reference Millán and Casanova20] and mixed infections [Reference Sobrinho21].
More specifically, the aim of this study was to evaluate the consequences of overlooking a part of the pathogen community in the explanatory power (EP) of statistical models designed for evaluating body-condition losses due to co-infection.
METHODS
Sampling procedure
In autumn 2008, 57 adult feral cats were live-trapped in Mallorca Island (Spain) with baited traps during a pest control campaign carried out to protect the endemic avifauna. Most of a cat's diet in the study area relies on rodent predation [Reference Millán22], hence we assumed that highly parasitized cats would show limited opportunities to hunt and hence the opportunity to compensate the cost of infection.
Once captured, cats were anaesthetized with a combination of ketamine (Imalgène®, Merial, France) plus xylazine (Rompun®, Bayer, Spain). Blood samples were collected from the cephalic vein and whole blood preserved in EDTA and sera were stored at −20°C until assayed. Subsequently, cats were humanely euthanatized with sodium thiopental (B. Braun Medical S.A., Spain). Only cats showing permanent teeth (aged >6 months) were used in this study. At the laboratory cats were necropsied in detail. Later, different species belonging to viruses, protozoa, helminths and arthropods were detected using specific laboratory methodologies that are summarized in Table 1. On the other hand, kidney fat reserves were used as a proxy for body condition in cats. Kidney fat reserves have already been used as a proxy for body reserves in other felids (e.g. European lynx, Lynx lynx [Reference Pulliainen, Lindgres and Tunkkari23]) and their statistical properties are well know [Reference Serrano24].
Table 1. Detection method, and associated pathology for the specific pathogen community for exploring the effects of multiple infections on body-condition losses in adult feral cats from Majorca Island, Spain

FIV, Feline immunodeficiency virus; FeLV, feline leukaemia virus; FPV, feline parvovirus; c-ELISA, competitive enzyme-linked immunosorbent assay; MAT, modified agglutination test; AI, active infection; C, contact.
* Joyeuxiella pasqualei, Diplopylidium acanthotetra, Dipylidium carracidoi, Taenia taeniaformis.
† Mostly Rhipicephalus sanguineus
‡ Mostly Ctenophalides felis. For FIV, the presence of antibodies indicates active infection; for Toxoplasma gondii cats showing high MAT titres (<1:200) were considered recently infected.
Statistical modelling
The effects of co-infection with viruses, protozoa, helminths and arthropods on cats' body condition were assessed by a Partial Least Squares Regression (PLS-R) approach. This statistical tool is an extension of multiple regression analysis. In the PLS-R, associations between explanatory and response variables are established with latent factors inferred through a mathematical model from variables measured directly. These latent factors are defined as linear combinations between predictor and response variables that maximize the explained variance in the dependent variables. During this iterative method the original multidimensionality is reduced to a lower number of orthogonal factors to detect the structure in the relationships between predictor variables and between these latent factors and the response variables. The extracted factors account for successively lower proportions of original variance [Reference Abbi and Salkind25].
PLS-R is probably the least restrictive of the multivariate techniques [Reference Haenlein and Kaplan26]. This flexibility allows it to be used when there are fewer observations than predictor variables, in case of multicollinearity and in case of studies of covariance in both explanatory and predictor variable groups [Reference Geladi and Kowalski27]. A comprehensive reading on the suitability of PLS-R in the fields of ecology and evolution can be found in Carrascal et al. [Reference Carrascal, Galván and Gordo28].
In our case, body condition (i.e. kidney fat) was the single explanatory variable while selected pathogen group combinations from the cats' pathogen community served as the predictor factor. Each pathogen group was defined by several species (Table 1). Furthermore, pathogen richness (e.g. the number of pathogen species actually observed per host) was also calculated and included as a part of the X component representing co-infection. Intensity of helminth infection was calculated according to Bush et al. [Reference Bush29].
Finally the EP for each of the candidate models, defined as the relative proportion of observed body-condition variability explained by the best PLS-R model, was also calculated. PLS-R was estimated with ‘plspm’ version 0.3·7 [Reference Sánchez and Trinchera30] of the Statistical package of R, version 3.0·1 [31].
RESULTS AND DISCUSSION
Only one cat was exclusively infected by a single group of parasites (helminths), confirming that multiple infections are the rule rather than the exception in wildlife [Reference Bordes and Morand32]. In fact, co-infections by only two parasite groups were rare (between 1·7% by arthropods and viruses, or 8·7% by helminths and protozoa), being 71·5% of individuals co-infected by ⩾3 parasite types (e.g. 31·5% by arthropods, helminths, protozoa and viruses, 19·0% by arthropods, helminths, and viruses, 12·3% by arthropods, helminths, and protozoa, and 8·7% by helminths, protozoa and viruses). The best two PLS-R models (see Table 2) provided a first component based on the combination of helminths, protozoa and viruses explaining 29·15% of body-condition variability of cats. Hence we assumed that this model had the maximum explanatory power (EP = 1). Such models suggested that cats in better condition were parasitized by few helminth and protozoa species (low species richness), i.e. lower Diplopylidium acanthotetra, Ancylostoma tubaeforme and Joyeuxiella pasqualei burdens, and were not infected by Toxoplasma gondii or with Leishmania infantum (Table 3, Fig. 1).
Table 2. Explanatory power (EP) of pathogen combinations belonging to different taxonomic groups for explaining the variations of fat reserves in free-roaming cats from Majorca Island, Spain

Models were ranked with respect to the percentage of observed body-condition variability (R 2Y) explained by a specific co-infection. The R 2X value indicates the percentage of variability of the X component explained by a specific co-infection of parasites belonging to one or different taxonomic groups.
Q 2 index (i.e. leave-one-out cross-validation); S indicates statistically significant Partial Least Squares Regression (PLS-R) model (α = 0·05); n.s. non-significant. EP indicates the percentage of explained variance with respect to the PLS-R model (i.e. helminths + protozoa + viruses) explaining the greatest proportion of observed body-condition variability (i.e. 29·15%).
Table 3. Predictor weights of the most parsimonious Partial Least Squares Regression (PLS-R) model explaining the effects of co-infection by several viruses, protozoa, helminth and arthropod species on body condition in 57 feral cats from Majorca Island, Spain

Predictor weights represent the contribution of each pathogen infection to the PLS-R x-axis. Predictor weights explaining more than 5% of the total variance in each response variable are shown in bold type.
It is interesting to note that helminths alone accounted for 76% of the EP (Table 2). This parasite group is normally acquired when cats consume an infected host or by kittens during nursing from an infected mother [Reference Millán and Casanova20]. The early infected animals are more likely to be clinically affected than cats infected at the adult stage, mainly because their immunological system is not fully developed. In fact, helminth-infected cats (especially young animals) can display diarrhoea, vomiting, permanent failure to grow and probably a reduced ability to store body reserves. However, while the influence of the whole helminth on body condition of cats is clear, Toxocara cati appears to be the most relevant species driving normal fat storage (Table 3, Fig. 1). In fact, T. cati is the cause of general failure to thrive in kittens harbouring moderate burdens, producing during its migration severe lesions in internal organs of adults [Reference Bowman33]. Cats parasitized by several parasite species harboured lower Toxocara loads, probably suggesting that few animals were able to deal with the effect of multiple infections in the case of Toxocara infection.
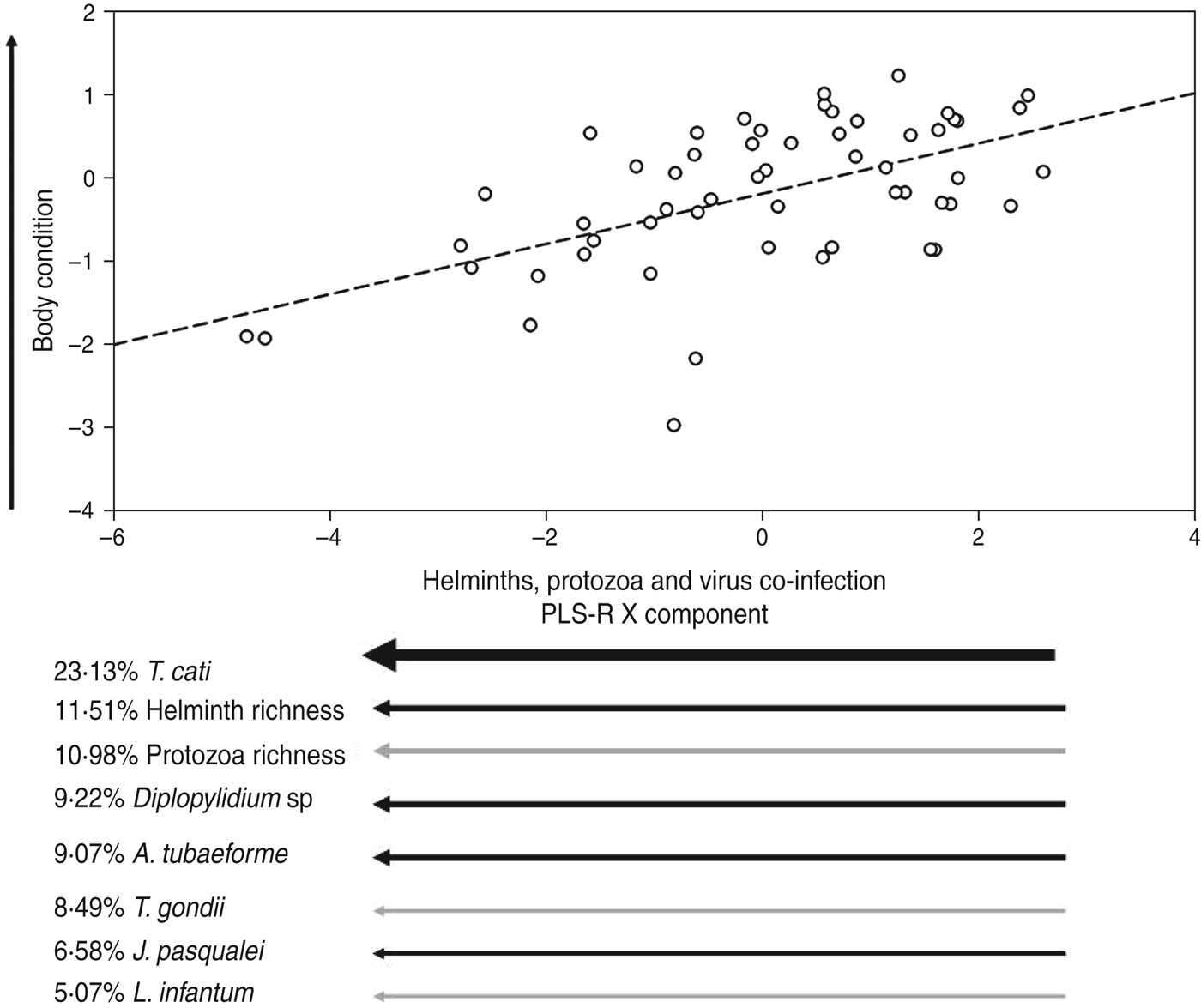
Fig. 1. Relationship between co-infecting helminths (black arrows) and protozoa (grey arrows) on a PLS-R component describing the body condition of adult feral cats. This plot represents the best PLS-R model shown in Table 2. Arrow direction indicates either an increase or a decrease of the component value, and arrow thickness directly indicates the weight of the component. Viruses explained <5% of the PLS-R X component and were therefore excluded from the plot.
On the other hand, both helminth and protozoa richness were included within the set of variables with substantial support underlining the role of protozoa infection (Toxoplasma and Leishmania) for normal body-condition storage. In addition, the exclusion of certain pathogen groups had a clear impact on the EP of models for explaining body-condition losses. In fact those including only one pathogen group lost between 24% and 94% of EP, depending on the pathogen group. For example, those models including helminths showed a slight reduction (<10%) in terms of EP with respect to the best model. Finally, we were surprised to observe the low impact of viral infections on body condition of cats. This could be explained by the fact that feline leukaemia virus and feline immunodeficiency virus infections can kill cats at the kitten or adult stage, and thus those infected animals we sampled may have shown some degree of resistance to these viral infections and hence a normal ability for fat storage.
We have shown here that neglecting some taxa of the host–parasite community diminishes the EP of models for exploring the cost of infection. The fact that both helminth and protozoa richness were included within the set of variables with substantial support, suggests the importance of considering as many pathogen species as possible in early models for studying the cost of infection. Helminths appeared to be the main group involved in explaining such variations in body condition, having been included in the models with the best EP. The high prevalence of this group of parasites and the fact that cats usually become infected at an early age [Reference Millán and Casanova20], suggests a relevant role for helminths in shaping the composition and impact of the remaining pathogen community. Evidence of this has been described for a broad range of host models [Reference Ezenwa and Jolles15] including humans [Reference Pullan and Brooker3], but still largely ignored in large-scale studies of co-infection [Reference Telfer8].
Our results support this idea and highlight a serious gap in the research directed to assess not only the impact of single infections but also that of co-infections on both animal and human health. From the community ecology point of view, outcomes of co-infection are difficult to generalize but particularly so when a part of the parasite community is neglected [Reference Pedersen and Fenton4]. Synergistic or antagonistic interactions between all types of pathogen combinations are, in fact, possible in natural conditions [Reference Cox34], and thus further research should take into account a broad representation of the whole pathogen community, especially those species interacting with the host from the early stages of development. This new view, based on considering pathogen communities rather than single relevant pathogens, will allow exploration of the interaction between such ‘host–parasite ecosystems’ and their natural environments [Reference Serrano35] providing perspectives for management and control of infectious diseases.
DECLARATION OF INTEREST
None.
ACKNOWLEDGEMENTS
The authors thank the Fundació Natura Parc for collaboration during fieldwork, Professor L. L. Vizcaino (UM) for laboratory analyses and Dr F. J. Márquez (UJAEN) for arthropod identification. This work was partially funded by Conselleria de Medi Ambient, Govern de les Illes Balears (contract no. 5893/2008). E. Serrano was supported by the Beatriu de Pinós programme and J. Millán by the Ramón y Cajal programme from the MICINN, Spain. Field work complied with current Spanish law.




