



1. Introduction
The Middle and Late Triassic were critical times in evolution. The global ecosystems recovered fully from the end-Permian mass extinction and started a full radiation both in the ocean and on land (Chen & Benton, Reference Chen and Benton2012; Sun et al. Reference Sun, Joachimski, Wignall, Yan, Chen, Jiang, Wang and Lai2012; Benton et al. Reference Benton, Zhang, Hu, Chen, Wen, Liu, Huang, Zhou, Xie and Tong2013, Reference Benton, Forth and Langer2014; Marshall, Reference Marshall2019). The marine realm saw the onset of the Mesozoic marine revolution, accompanied by an increase in predation pressure in an increasingly complex marine food web (Vermeij, Reference Vermeij1977; Harper et al. Reference Harper, Forsythe and Palmer1998; Benton et al. Reference Benton, Forth and Langer2014; Kelley & Pyenson, Reference Kelley and Pyenson2015; Tackett, Reference Tackett2016). The environmental, biotic and climatic changes have been intensively studied in this interval (e.g. Mutti & Weissert, Reference Mutti and Weissert1995; Wang et al. Reference Wang, Bachmann, Hagdorn, Sander, Cuny, Chen, Wang, Chen, Cheng and Meng2008; Stefani et al. Reference Stefani, Furin and Gianolla2010; Benton et al. Reference Benton, Zhang, Hu, Chen, Wen, Liu, Huang, Zhou, Xie and Tong2013; Trotter et al. Reference Trotter, Williams, Nicora, Mazza and Rigo2015; Tanner, Reference Tanner2018; Sun et al. Reference Sun, Orchard, Kocsis and Joachimski2020). However, lining up key evolutionary events with palaeoenvironmental changes at a global scale requires a robust biostratigraphic framework and high precision supra-regional correlation, which remain challenging in many cases (e.g. Kozur, Reference Kozur2003; Mundil et al. Reference Mundil, Palfy, Renne, Brack and Lucas2010; Chen et al. Reference Chen, Krystyn, Orchard, Lai and Richoz2015; Shen & Zhang, Reference Shen and Zhang2017; Rigo et al. Reference Rigo, Mazza, Karádi, Nicora and Tanner2018; Tong et al. Reference Tong, Chu, Liang, Shu, Song, Song, Song and Wu2019; Gradstein et al. Reference Gradstein, Ogg, Schmitz and Ogg2020).
The Global Stratotype Section and Point for the base of the Carnian Stage is defined by the first appearance datum (FAD) of the ammonoid Daxatina canadensis at Prati di Stuores in NE Italy (Mietto et al. Reference Mietto, Manfrin, Preto, Rigo, Roghi, Furin, Gianolla, Posenato, Muttoni and Nicora2012). Correlations between the western Tethys and North America for the Ladinian–Carnian boundary (L-CB) beds are well resolved (Mietto & Manfrin, Reference Mietto and Manfrin1995; Orchard & Tozer, Reference Orchard and Tozer1997; Gallet et al. Reference Gallet, Krystyn and Besse1998; Balini et al. Reference Balini, Germani, Nicora and Rizzi2000, Reference Balini, Lucas, Jenks, Spielmann and Lucas2010; Mietto et al. Reference Mietto, Andreetta, Broglio, Buratti, Cirilli, De Zanche, Furin, Gianolla, Manfrin, Muttoni, Neri, Nicora, Posenato, Preto, Rigo, Roghi and Spötl2007, Reference Mietto, Manfrin, Preto, Rigo, Roghi, Furin, Gianolla, Posenato, Muttoni and Nicora2012; Orchard, Reference Orchard, Lucas and Spielmann2007). However, correlating time-equivalent beds in the eastern and western Tethys has proven challenging. This is largely because the biostratigraphic definition of the L-CB in South China remains ambiguous owing to paucities of the ammonoids Daxatina canadensis and associated Trachyceras species (Tong et al. Reference Tong, Chu, Liang, Shu, Song, Song, Song and Wu2019). Several options were proposed, and some led to discrepant solutions (e.g. Xu et al. Reference Xu, Niu and Chen2003; Sun et al. Reference Sun, Hao and Jiang2005; Zou et al. Reference Zou, Balini, Jiang, Tintori, Sun and Sun2015).
A common practice is to define the L-CB using the FAD of ‘Paragondolella’ (P.) polygnathiformis, which was ratified as the secondary auxiliary marker for the basal Carnian at Prati di Stuores (Yang et al. Reference Yang, Liu and Zhang1995; Sun et al. Reference Sun, Hao and Jiang2005; Mietto et al. Reference Mietto, Manfrin, Preto, Rigo, Roghi, Furin, Gianolla, Posenato, Muttoni and Nicora2012; Muttoni et al. Reference Muttoni, Mazza, Mosher, Katz, Kent and Balini2014; Lehrmann et al. Reference Lehrmann, Stepchinski, Altiner, Orchard, Montgomery, Enos, Ellwood, Bowring, Ramezani and Wang2015). However, the FAD of P. polygnathiformis is difficult to define in practice, and large ontogenetic variation within the species hinders the resolution of this issue further (e.g. Koike, Reference Koike1982; Koike et al. Reference Koike, Kodachi, Matsuno and Baba1991; Orchard & Balini, Reference Orchard, Balini, Lucas and Spielmann2007; Orchard, Reference Orchard and Lucas2010; Chen & Lukeneder, Reference Chen and Lukeneder2017; Jiang et al. Reference Jiang, Chen and Liu2018). Therefore, additional markers for the L-CB are in urgent need.
Subdividing Carnian strata is a difficult task. Conodont zonation, especially in the lower Carnian (Julian), varies significantly in different regions (Koike, Reference Koike, Koike and Igo1979; Kozur, Reference Kozur1980 a, Reference Kozur2003; Budurov et al. Reference Budurov, Gupta, Sudar and Buryi1985; Orchard & Tozer, Reference Orchard and Tozer1997; Chen et al. Reference Chen, Krystyn, Orchard, Lai and Richoz2015; Rigo et al. Reference Rigo, Mazza, Karádi, Nicora and Tanner2018; Yamashita et al. Reference Yamashita, Kato, Onoue and Suzuki2018). This is partly due to few studies being carried out and partly due to an increase in endemism in conodonts since late Middle Triassic time (e.g. Budurov et al. Reference Budurov, Gupta, Sudar and Buryi1985; Chen et al. Reference Chen, Krystyn, Orchard, Lai and Richoz2015). In North America, two conodont zones, namely the Neospathodus newpassensis Zone and the P. polygnathiformis Zone, were initially established for the lower Carnian in Nevada and British Columbia (Sweet et al. Reference Sweet, Mosher, Clark, Collinson, Hasenmueller, Stweet and Bergstrom1971). The Metapolygnathus (Me.) polygnathiformis Zone and the P. inclinata–Mosherella assemblage zone (AZ) were summarized for the lower Carnian in the Western Canada Sedimentary Basin (Orchard & Tozer, Reference Orchard and Tozer1997). Later, the P. inclinata Zone (which consisted of the P. sulcata Subzone and the Me. acuminata Subzone), the Me. intermedia Zone and the Me. tadpole Zone were recognized in British Columbia (Orchard, Reference Orchard, Lucas and Spielmann2007). In the western Tethys, three conodont zones, namely, the Carinella diebeli AZ, the Gladigondolella (G.) tethydis AZ and the Gondolella polygnathiformis AZ were established for the upper Ladinian to the lower Carnian in the Salzkammergut region of the North Calcareous Alps (Krystyn, Reference Krystyn1980). The P. foliata range zone (RZ) and the P. polygnathiformis interval zone (IZ) were established based on collections from the Alps, Dinarides, Balkans and Himalayas (Budurov & Sudar, Reference Budurov and Sudar1990). Later, five conodont zones, namely, the Budurovignathus (B.) mostleri IZ, the Me. tadpole IZ, the Me. auriformis IZ, the Me. carnica RZ and the G. tethydis IZ were recognized in the North Calcareous Alps and southwestern Turkey in the lower Carnian (Gallet et al. Reference Gallet, Besse, Krystyn, Théveniaut and Marcoux1994). Rigo et al. (Reference Rigo, Mazza, Karádi, Nicora and Tanner2018) proposed a new lower Carnian conodont scheme for the western Tethys, consisting of the P. polygnathiformis IZ, the Mazzaella (Ma.) carnica IZ and the P. praelindae IZ. In the Panthalassa Ocean, two zones, namely, the Neogondolella (N.) foliata Zone and N. polygnathiformis Zone, were summarized for the L-CB interval in Japan (Koike, Reference Koike, Koike and Igo1979). In central Japan, only the P. tadpole IZ was initially established for the lower Carnian (Yamashita et al. Reference Yamashita, Kato, Onoue and Suzuki2018). This was later revised to the Ma. carnica Zone and the P. praelindae Zone in the same section (Tomimatsu et al. Reference Tomimatsu, Nozaki, Sato, Takaya, Kimura, Chang, Naraoka, Rigo and Onoue2021). In addition, the Quadralella (Q.) tadpole – G. malayensis AZ was established in Kamura in southern Japan for the lower Carnian (Zhang et al. Reference Zhang, Orchard, Algeo, Chen, Lyu, Zhao, Kaiho, Ma and Liu2019).
Much progress has been made in the Carnian conodont biostratigraphy in South China (e.g. Wang et al. Reference Wang, Kang and Wang1998, Reference Wang, Bachmann, Hagdorn, Sander, Cuny, Chen, Wang, Chen, Cheng and Meng2008; Yang et al. Reference Yang, Hao and Jiang2002; Sun, Z. Y. et al. Reference Sun, Hao and Jiang2005, Reference Sun, Jiang, Ji and Hao2016; Jin et al. Reference Jin, Shi, Rigo, Franceschi and Preto2018). Early studies only recognized two conodont zones, the B. diebeli Zone and the P. polygnathiformis Zone, for the entire Carnian (Yao, Reference Yao1987; Lai & Mei, Reference Lai and Mei2000). The N. polygnathiformis – N. maantangensis AZ, N. polygnathiformis – N. tadpole AZ and N. polygnathiformis Zone were established by Yang et al. (Reference Yang, Liu and Zhang1995) in Guizhou. Yang et al. (Reference Yang, Hao, Wang, Yao, Ezaki, Hao and Wang1999) noticed that conodont assemblages vary with depositional settings and established the B. diebeli – P. sp. A AZ, the P. polygnathiformis Zone and the ‘Epigondolella’ nodosa Zone for basinal environments and the P. polygnathiformis – P. maantangensis AZ, the P. polygnathiformis – P. tadpole AZ and the P. polygnathiformis Zone for platform environments (Yang et al. Reference Yang, Hao, Wang, Yao, Ezaki, Hao and Wang1999; Wang & Wang, Reference Wang and Wang2016). Y. D. Sun et al. (Reference Sun, Wignall, Joachimski, Bond, Grasby, Lai, Wang, Zhang and Sun2016) identified the P. foliata – Q. polygnathiformis AZ, the Q. polygnathiformis noah AZ and the Q. ex gr. carpathica AZ in a carbonate ramp section in Guizhou. Zhang et al. (Reference Zhang, Sun and Lai2018 a) summarized eight conodont zones in southwestern China: the P. foliata Zone, the Q. polygnathiformis Zone, the Q. tadpole Zone, the Q. praelindae Zone, the Q. aff. auriformis Zone, the Q. robusta Zone, the Q. noah Zone and the Q. ex gr. carpathica Zone. Jiang et al. (Reference Jiang, Yuan, Chen, Ogg and Yan2019) identified the Ma. carnica RZ in northern Sichuan.
The subdivision of the Middle and Upper Triassic strata into substages or finer levels is a longstanding conundrum in South China. To address this issue, we carried out a study in a newly excavated section in eastern Yunnan Province. Here, we define the L-CB in the Zhuganpo Formation (Fm) by using the first occurrence (FO) of Q. polygnathiformis and the FO of Q. intermedia, providing a practical solution for the L-CB definition in the region. A new conodont biostratigraphic framework is established, and the zones can be well correlated to other regions. Our study also reveals that the conodont fauna from southwestern China shows mixed morphological features compared to their North American and western Tethyan counterparts.
2. Geological setting
The South China Block has been a classic area for Triassic research. It was located in the northern low latitudes in the eastern Palaeo-Tethys during Late Triassic time and consisted of the Yangtze Platform, the Cathaysian Platform and the Nanpanjiang Basin (Fig. 1; Yin & Peng, Reference Yin and Peng2000). Marine carbonates with diverse facies were extensively developed in the region during Early Triassic time. The carbonate platform continued to shrink toward Late Triassic time, partly due to the Indosinian Orogeny. Most Middle to Upper Triassic carbonate successions were deposited on the southwestern margin of the Yangtze Platform and sparsely in the Nanpanjiang Basin. The studied Yize section is located near Changdi town in Luoping city, Yunnan Province (Fig. 1), palaeogeographically belonging to the Upper Yangtze Platform.

Fig. 1. The palaeogeographical and present-day location maps of the study section. (a) Palaeogeographical map during Carnian time (modified from Sun et al. Reference Sun, Richoz, Krystyn, Zhang and Joachimski2019). (b, c) Present-day location maps of the Yize section.
The Yize section was freshly exposed along a hillside quarry. The measured section is 95 m in thickness and consists of the Zhuganpo and the Wayao (also known as Xiaowa) formations. The measured Zhuganpo Fm is ∼90 m in thickness and is dominated by bioturbated carbonates. The unit is characterized by three distinct lithological associations. They are, in ascending order, the fine-laminated limestone member, the cherty limestone member and the nodular limestone member. The overlying Wayao Fm is ∼5 m in thickness, comprising thin- to medium-bedded sandstones and mudstones (Figs 2, 3).

Fig. 2. Field photographs of the study section at Yize, Yunnan Province, South China. (a) Overview of the studied section, showing the outcrop of the Zhuganpo and the Wayao formations. (b) Photo showing the fine-laminated limestones in the lower Zhuganpo Fm. (c, d) Photos showing the cherty limestone member of the Zhuganpo Fm. (e) Photo showing the thin- to medium-bedded sandstones and mudstones of the lower Wayao Fm.

Fig. 3. Log of the Yize section, showing sample positions, conodont occurrences and zonation.
3. Materials and methods
The study section was logged in detail in the field and systematically sampled for conodonts with a resolution of ∼1–3 m. Forty-three carbonate samples, each weighing 7–9 kg, were collected. All samples were crushed into 1–2 cm3 rock chips and dissolved in diluted acetic acid (∼8 vol. %). The undissolved residuals were wet-sieved and dried at room temperature. The heavy fractions were separated using a lithium and sodium heteropolytungstate heavy liquid (density 2.80 g cm−3). Conodont elements were picked out under a binocular microscope. A total of 1234 conodont (989 P1 and 245 ramiform) elements were recovered from 37 samples. Selected specimens were imaged using a scanning electron microscope (SEM) at the China University of Geosciences (Wuhan).
4. Conodont biostratigraphy
Five conodont zones are established and described below in ascending order. Reported conodont taxa and their ranges are shown in Figure 3. Note that some genera and species are revised from their original binomial nomenclature and reassigned in accordance with the latest taxonomic studies and rules of the International Code of Zoological Nomenclature (e.g. Orchard, Reference Orchard, Lucas and Spielmann2007, Reference Orchard, Tanner, Spielmann and Lucas2013; Chen et al. Reference Chen, Krystyn, Orchard, Lai and Richoz2015; Kiliç et al. Reference Kiliç, Plasencia, Ishida and Hirsch2015, Reference Kiliç, Plasencia, Guex, Hirsch and Montenari2017; Chen & Lukeneder, Reference Chen and Lukeneder2017). For a better description of the stratigraphic context, we use stage and substage nomenclature in the following sections.
4.a. Paragondolella inclinata Zone
Lower limit: not defined.
Upper limit: FO of Q. polygnathiformis.
Associated taxa: P. inclinata, Q. acuminata (= Me. acuminatus in Orchard, Reference Orchard, Lucas and Spielmann2007) and Q. lobata (= Me. lobatus in Orchard, Reference Orchard, Lucas and Spielmann2007).
The P. inclinata Zone ranges from the 13.5 m to the 29.4 m level at Yize. This zone represents the last Ladinian conodont zone in South China and British Columbia (Orchard & Tozer, Reference Orchard and Tozer1997; Orchard, Reference Orchard, Lucas and Spielmann2007; Lehrmann et al. Reference Lehrmann, Stepchinski, Altiner, Orchard, Montgomery, Enos, Ellwood, Bowring, Ramezani and Wang2015). The zonal species is cosmopolitan ranging from the lower Longobardian to the upper Julian (Gallet et al. Reference Gallet, Krystyn and Besse1998; Orchard, Reference Orchard, Lucas and Spielmann2007; Rigo et al. Reference Rigo, Preto, Roghi, Tateo and Mietto2007; Lein et al. Reference Lein, Krystyn, Sylvain and Henry2012; Chen et al. Reference Chen, Krystyn, Orchard, Lai and Richoz2015; Zhang et al. Reference Zhang, Sun, Lai, Joachimski and Wignall2017). Morphologically, the species represents a transitional form from P. excelsa and P. foliata (Kovács, Reference Kovács1983) (Fig. 4).

Fig. 4. SEM photographs of conodonts from the Ladinian to Carnian strata at Yize. Scale bar = 200 μm. a – upper view; b – lateral view; c – lower view. 1 – Q. polygnathiformis (Budurov & Stefanov, Reference Budurov and Stefanov1965), YZ-(-65)-001; 2 – Q. intermedia (Orchard, Reference Orchard, Lucas and Spielmann2007), YZ-(-65)-014; 3 – Q. polygnathiformis (Budurov & Stefanov, Reference Budurov and Stefanov1965), YZ-12-1-014; 4 – Q. jiangyouensis (Wang & Dai, Reference Wang and Dai1981), YZ-25-003; 5 – Q. polygnathiformis (Budurov & Stefanov, Reference Budurov and Stefanov1965), YZ-(-65)-015; 6 – Q. polygnathiformis (Budurov & Stefanov, Reference Budurov and Stefanov1965), YZ-88-1-36; 7 – Q. aff. polygnathiformis magna (Igo, Reference Igo1989), YZ-18-001; 8 – Q. polygnathiformis (Budurov & Stefanov, Reference Budurov and Stefanov1965), YZ-12-1-013; 9 – Q. aff. polygnathiformis magna (Igo, Reference Igo1989), YZ-12-2-001; 10 – Q. maantangensis (Dai & Tian in Tian et al. Reference Tian, Dai and Tian1983), YZ-25-001; 11 – P. inclinata (Kovács, Reference Kovács1983), YZ-25-002; 12 – P. inclinata (Kovács, Reference Kovács1983), YZ-28-001; 13 – K. praeangustus (Kozur, Miräuta & Mock in Kozur, Reference Kozur1980 b), YZ-42-020; 14 – Q. polygnathiformis (Budurov & Stefanov, Reference Budurov and Stefanov1965), YZ-42-026; 15 – Q. intermedia (Orchard, Reference Orchard, Lucas and Spielmann2007), YZ-79-017; 16 – Q. intermedia (Orchard, Reference Orchard, Lucas and Spielmann2007), YZ-63-005; 17 – Q. polygnathiformis (Budurov & Stefanov, Reference Budurov and Stefanov1965), YZ-63-007; 18 – Q. intermedia (Orchard, Reference Orchard, Lucas and Spielmann2007), YZ-79-009; 19 – Q. intermedia (Orchard, Reference Orchard, Lucas and Spielmann2007), YZ-95-1-003; 20 – Q. sp., YZ-63-008; 21 – Q. jiangyouensis (Wang & Dai, Reference Wang and Dai1981), YZ-79-026; 22 – Q. tadpole (Hayashi, Reference Hayashi1968), YZ-95-1-006; 23 – Q. robusta Zhang, Sun & Lai in Zhang et al. Reference Zhang, Sun, Lai and Wignall2018 b, YZ-95-2-022; 24 – Q. polygnathiformis (Budurov & Stefanov, Reference Budurov and Stefanov1965), YZ-95-1-005.
In British Columbia, Q. acuminata and Q. lobata were identified in the P. inclinata Zone and the Q. intermedia Zone, respectively. Both species typically represent the Longobardian to Julian elements (Orchard, Reference Orchard, Lucas and Spielmann2007). Q. acuminata and Q. lobata are also known in the Zhuganpo Fm from Guizhou and Yunnan in southwestern China as well as in Japan (Zhang et al. Reference Zhang, Sun, Lai, Joachimski and Wignall2017, Reference Zhang, Sun, Lai and Wignall2018 b, Reference Zhang, Orchard, Algeo, Chen, Lyu, Zhao, Kaiho, Ma and Liu2019).
4.b. Quadralella polygnathiformis Zone
Lower limit: FO of Q. polygnathiformis.
Upper limit: FO of Q. praelindae.
Associated taxa: P. foliata, P. inclinata, Q. acuminata, Q. intermedia (= Me. intermedius in Orchard, Reference Orchard, Lucas and Spielmann2007), Q. jiangyouensis, Q. langdaiensis, Q. lobata, Q. maantangensis, Q. aff. polygnathiformis magna, Q. spp., Q. shijiangjunensis, Q. tadpole, Q. wanlanensis and Q. yongyueensis.
The Q. polygnathiformis Zone ranges from the 29.4 m to the 47.1 m level at Yize. This zone was widely accepted as the first Carnian conodont zone (e.g. Orchard & Tozer, Reference Orchard and Tozer1997; Kozur, Reference Kozur2003; Rigo et al. Reference Rigo, Mazza, Karádi, Nicora and Tanner2018; Tong et al. Reference Tong, Chu, Liang, Shu, Song, Song, Song and Wu2019). The FAD of Q. polygnathiformis is the secondary marker to define the base of the Carnian (Mietto et al. Reference Mietto, Manfrin, Preto, Rigo, Roghi, Furin, Gianolla, Posenato, Muttoni and Nicora2012). Q. polygnathiformis ranges typically from the lower Julian to the lower Tuvalian and may extend to the upper Tuvalian (e.g. Gallet et al. Reference Gallet, Besse, Krystyn, Théveniaut and Marcoux1994; Rigo et al. Reference Rigo, Preto, Roghi, Tateo and Mietto2007, Reference Rigo, Mazza, Karádi, Nicora and Tanner2018; Muttoni et al. Reference Muttoni, Mazza, Mosher, Katz, Kent and Balini2014; Chen et al. Reference Chen, Krystyn, Orchard, Lai and Richoz2015). Some studies implied that Q. polygnathiformis might appear in older strata below the FAD of Daxatina canadensis (Krystyn et al. Reference Krystyn, Balini and Nicora2004; Orchard, Reference Orchard, Lucas and Spielmann2007, Reference Orchard and Lucas2010; Mietto et al. Reference Mietto, Manfrin, Preto, Rigo, Roghi, Furin, Gianolla, Posenato, Muttoni and Nicora2012). The essential feature of this species is the geniculation(s)/abrupt step(s) at the anterior margins, which differs from Ladinian holdovers. However, Q. polygnathiformis may show large morphological variations in the posterior platform during different ontogenetic stages (e.g. Orchard & Balini, Reference Orchard, Balini, Lucas and Spielmann2007; Chen & Lukeneder, Reference Chen and Lukeneder2017) (Fig. 5).

Fig. 5. SEM photographs of conodonts from Ladinian to Carnian strata at Yize. Scale bar = 200 μm. a – upper view; b – lateral view; c – lower view. 1 – Q. polygnathiformis (Budurov & Stefanov, Reference Budurov and Stefanov1965), YZ-95-1-029; 2 – K. praeangustus (Kozur, Miräuta & Mock in Kozur, Reference Kozur1980 b), YZ-95-1-030; 3 – Q. jiangyouensis (Wang & Dai, Reference Wang and Dai1981), YZ-95-1-031; 4 – K. praeangustus (Kozur, Miräuta & Mock in Kozur, Reference Kozur1980 b), YZ-95-1-032; 5 – Q. polygnathiformis (Budurov & Stefanov, Reference Budurov and Stefanov1965), YZ-95-1-033; 6 – Q. intermedia (Orchard, Reference Orchard, Lucas and Spielmann2007), YZ-95-1-034; 7 – Q. robusta Zhang, Sun & Lai in Zhang et al. Reference Zhang, Sun, Lai and Wignall2018 b, YZ-95-1-036; 8 – P. inclinata (Kovács, Reference Kovács1983), YZ-95-1-044; 9 – Q. jiangyouensis (Wang & Dai, Reference Wang and Dai1981), YZ-95-1-017; 10 – Q. tadpole (Hayashi, Reference Hayashi1968), YZ-95-2-046; 11 – Q. intermedia (Orchard, Reference Orchard, Lucas and Spielmann2007), YZ-95-1-061; 12 – Q. aff. zonneveldi (Orchard, Reference Orchard, Lucas and Spielmann2007), YZ-95-1-066; 13 – P. foliata Budurov, Reference Budurov1975, YZ-95-1-070; 14 – Q. tadpole (Hayashi, Reference Hayashi1968), YZ-95-2-019; 15 – Q. praelindae (Kozur, Reference Kozur2003), YZ-95-1-015; 16 – Q. praelindae (Kozur, Reference Kozur2003), YZ-95-1-009; 17 – Q. intermedia (Orchard, Reference Orchard, Lucas and Spielmann2007), YZ-95-2-012; 18 – Q. polygnathiformis (Budurov & Stefanov, Reference Budurov and Stefanov1965), YZ-95-2-016; 19 – Q. polygnathiformis (Budurov & Stefanov, Reference Budurov and Stefanov1965), YZ-95-2-024; 20 – Q. robusta Zhang, Sun & Lai in Zhang et al. Reference Zhang, Sun, Lai and Wignall2018 b, YZ-95-2-045; 21 – Q. robusta Zhang, Sun & Lai in Zhang et al. Reference Zhang, Sun, Lai and Wignall2018 b, YZ-95-2-047; 22 – Q. jiangyouensis (Wang & Dai, Reference Wang and Dai1981), YZ-95-2-049; 23 – K. praeangustus (Kozur, Miräuta & Mock in Kozur, Reference Kozur1980 b), YZ-95-2-051; 24 – K. praeangustus (Kozur, Miräuta & Mock in Kozur, Reference Kozur1980 b), YZ-95-2-052; 25 – K. praeangustus (Kozur, Miräuta & Mock in Kozur, Reference Kozur1980 b), YZ-95-2-053; 26 – P. inclinata (Kovács, Reference Kovács1983), YZ-99-025; 27 – P. inclinata (Kovács, Reference Kovács1983), YZ-12-2-002.
P. foliata is also a cosmopolitan species and differs from P. inclinata by having a straight basal edge before the basal cavity (Koike, Reference Koike1982; Kovács, Reference Kovács1983; Muttoni et al. Reference Muttoni, Mazza, Mosher, Katz, Kent and Balini2014; Zhang et al. Reference Zhang, Sun, Lai, Joachimski and Wignall2017). The species ranges from the Longobardian 3 to the Tuvalian 1 (Lein et al. Reference Lein, Krystyn, Sylvain and Henry2012; Muttoni et al. Reference Muttoni, Mazza, Mosher, Katz, Kent and Balini2014), but is most common in the Julian (Rigo et al. Reference Rigo, Preto, Roghi, Tateo and Mietto2007; Zhang et al. Reference Zhang, Sun, Lai, Joachimski and Wignall2017).
Q. intermedia was first identified in the Frankites sutherlandi ammonoid zone in British Columbia (Orchard, Reference Orchard, Lucas and Spielmann2007) and has been reported in Nevada and southwestern China (Orchard, Reference Orchard, Lucas and Spielmann2007; Orchard & Balini, Reference Orchard, Balini, Lucas and Spielmann2007; Zhang et al. Reference Zhang, Sun, Lai, Joachimski and Wignall2017, Reference Zhang, Sun, Lai and Wignall2018 b). The FO of Q. intermedia has been used as an effective marker for the base of the Carnian in British Columbia because of its co-occurrence with Daxatina canadensis and its unambiguous evolution lineage (Orchard, Reference Orchard, Lucas and Spielmann2007). This species is thought to derive from Q. acuminata through further reduction in the anterior platform (Orchard, Reference Orchard, Lucas and Spielmann2007). Reduction in the anterior platform and a narrowly rounded posterior end are key diagnostic features of Q. intermedia that differentiate the species from Q. polygnathiformis (Orchard, Reference Orchard, Lucas and Spielmann2007; Orchard & Balini, Reference Orchard, Balini, Lucas and Spielmann2007).
Q. tadpole is characterized by its tadpole-shaped platform and was widely reported in the Tethys, North America and Panthalassa (e.g. Hayashi, Reference Hayashi1968; Gallet et al. Reference Gallet, Besse, Krystyn, Théveniaut and Marcoux1994; Orchard, Reference Orchard, Lucas and Spielmann2007; Mietto et al. Reference Mietto, Manfrin, Preto, Rigo, Roghi, Furin, Gianolla, Posenato, Muttoni and Nicora2012; Zhang et al. Reference Zhang, Sun, Lai, Joachimski and Wignall2017, Reference Zhang, Orchard, Algeo, Chen, Lyu, Zhao, Kaiho, Ma and Liu2019). This species commonly occurred in the Julian and disappeared in the Tuvalian (Rigo et al. Reference Rigo, Preto, Roghi, Tateo and Mietto2007; Chen et al. Reference Chen, Krystyn, Orchard, Lai and Richoz2015; Kiliç et al. Reference Kiliç, Plasencia, Guex, Hirsch and Montenari2017; Tomimatsu et al. Reference Tomimatsu, Nozaki, Sato, Takaya, Kimura, Chang, Naraoka, Rigo and Onoue2021) (Fig. 6).

Fig. 6. SEM photographs of conodonts from Ladinian to Carnian strata at Yize. Scale bar = 200 μm. a – upper view; b – lateral view; c – lower view. 1 – Q. intermedia (Orchard, Reference Orchard, Lucas and Spielmann2007), YZ-95-1-004; 2 – Q. polygnathiformis (Budurov & Stefanov, Reference Budurov and Stefanov1965), YZ-88-1-17; 3 – Q. polygnathiformis (Budurov & Stefanov, Reference Budurov and Stefanov1965), YZ-88-1-18; 4 – Q. polygnathiformis (Budurov & Stefanov, Reference Budurov and Stefanov1965), YZ-88-1-19; 5 – Q. praelindae (Kozur, Reference Kozur2003), YZ-88-1-08; 6 – Q. intermedia (Orchard, Reference Orchard, Lucas and Spielmann2007), YZ-88-1-01; 7 – Q. tadpole (Hayashi, Reference Hayashi1968), YZ-88-1-04; 8 – Q. polygnathiformis (Budurov & Stefanov, Reference Budurov and Stefanov1965), YZ-88-1-05; 9 – Q. polygnathiformis (Budurov & Stefanov, Reference Budurov and Stefanov1965), YZ-88-1-07; 10 – Q. praelindae (Kozur, Reference Kozur2003), YZ-88-1-12; 11 – Q. tadpole (Hayashi, Reference Hayashi1968), YZ-88-1-16; 12 – Q. polygnathiformis (Budurov & Stefanov, Reference Budurov and Stefanov1965), YZ-88-1-14; 13 – Q. praelindae (Kozur, Reference Kozur2003), YZ-88-2-03; 14 – Q. polygnathiformis (Budurov & Stefanov, Reference Budurov and Stefanov1965), YZ-88-1-15; 15 – Q. intermedia (Orchard, Reference Orchard, Lucas and Spielmann2007), YZ-88-2-16; 16 – Q. intermedia (Orchard, Reference Orchard, Lucas and Spielmann2007), YZ-88-2-21; 17 – Q. sp., YZ-95-2-057; 18 – Q. praelindae (Kozur, Reference Kozur2003), YZ-88-1-09; 19 – Q. intermedia (Orchard, Reference Orchard, Lucas and Spielmann2007), YZ-88-2-22; 20 – Q. acuminata (Orchard, Reference Orchard, Lucas and Spielmann2007), YZ-(-17)2-001; 21 – P. inclinata (Kovács, Reference Kovács1983), YZ-(-17)2-002; 22 – Q. polygnathiformis (Budurov & Stefanov, Reference Budurov and Stefanov1965), YZ-(-65)-i001; 23 – Q. acuminata (Orchard, Reference Orchard, Lucas and Spielmann2007), YZ-(-65)-004; 24 – Q. intermedia (Orchard, Reference Orchard, Lucas and Spielmann2007), YZ-(-65)-005; 25 – Q. intermedia (Orchard, Reference Orchard, Lucas and Spielmann2007), YZ-(-65)-i005; 26 – P. foliata Budurov, Reference Budurov1975, YZ-(-65)-012; 27 – P. inclinata (Kovács, Reference Kovács1983), YZ-(-17)2-003.
Q. jiangyouensis, Q. langdaiensis, Q. maantangensis, Q. shijiangjunensis and Q. yongyueensis are only reported from South China to date (Wang & Dai, Reference Wang and Dai1981; Tian et al. Reference Tian, Dai and Tian1983; Yang et al. Reference Yang, Hao and Jiang2002; Sun, Reference Sun2006; Sun, Y. D. et al. Reference Sun, Wignall, Joachimski, Bond, Grasby, Lai, Wang, Zhang and Sun2016; Zhang et al. Reference Zhang, Sun, Lai, Joachimski and Wignall2017, Reference Zhang, Sun, Lai and Wignall2018 b). Q. aff. maantangensis was reported in central Japan (Tomimatsu et al. Reference Tomimatsu, Nozaki, Sato, Takaya, Kimura, Chang, Naraoka, Rigo and Onoue2021).
Q. polygnathiformis magna was first established in the Carnian in Japan (Igo, Reference Igo1989). The reported Q. aff. polygnathiformis magna in the Yize section is similar to Q. polygnathiformis magna in its bifurcated keel end but differs in its platform outline (Igo, Reference Igo1989). Q. aff. polygnathiformis magna is regionally common, also known from the Zhuganpo Fm in Yunnan Province (Zhang et al. Reference Zhang, Sun, Lai and Wignall2018 b). Q. wanlanensis was first established in the Julian in southwestern China (Zhang et al. Reference Zhang, Sun, Lai, Joachimski and Wignall2017, Reference Zhang, Sun, Lai and Wignall2018 b) and was later also recognized in central Japan (Tomimatsu et al. Reference Tomimatsu, Nozaki, Sato, Takaya, Kimura, Chang, Naraoka, Rigo and Onoue2021), suggesting the species may be geographically widespread.
4.c. Quadralella praelindae Zone
Lower limit: FO of Q. praelindae.
Upper limit: FO of Q. auriformis.
Associated taxa: G. malayensis, G. tethydis, Kraussodontus (K.) praeangustus, P. inclinata, Q. acuminata, Q. intermedia, Q. jiangyouensis, Q. lobata, Q. polygnathiformis, Q. aff. polygnathiformis magna, Q. shijiangjunensis, Q. spp., Q. tadpole, Q. wanlanensis, Q. wayaoensis, Q. yongyueensis and Q. aff. zonneveldi.
The Q. praelindae Zone ranges from the 47.1 m to the 67 m level. Q. praelindae was first identified in the lower Tuvalian at Silická Brezová, Slovakia (Kozur, Reference Kozur2003; Channell et al. Reference Channell, Kozur, Sievers, Mock, Aubrecht and Sykora2003). Later, the species was reported in the Julian strata (Rigo et al. Reference Rigo, Preto, Roghi, Tateo and Mietto2007; Sun, Z. Y. et al. Reference Sun, Wignall, Joachimski, Bond, Grasby, Lai, Wang, Zhang and Sun2016; Zhang et al. Reference Zhang, Sun, Lai and Wignall2018 b) (Fig. 7).
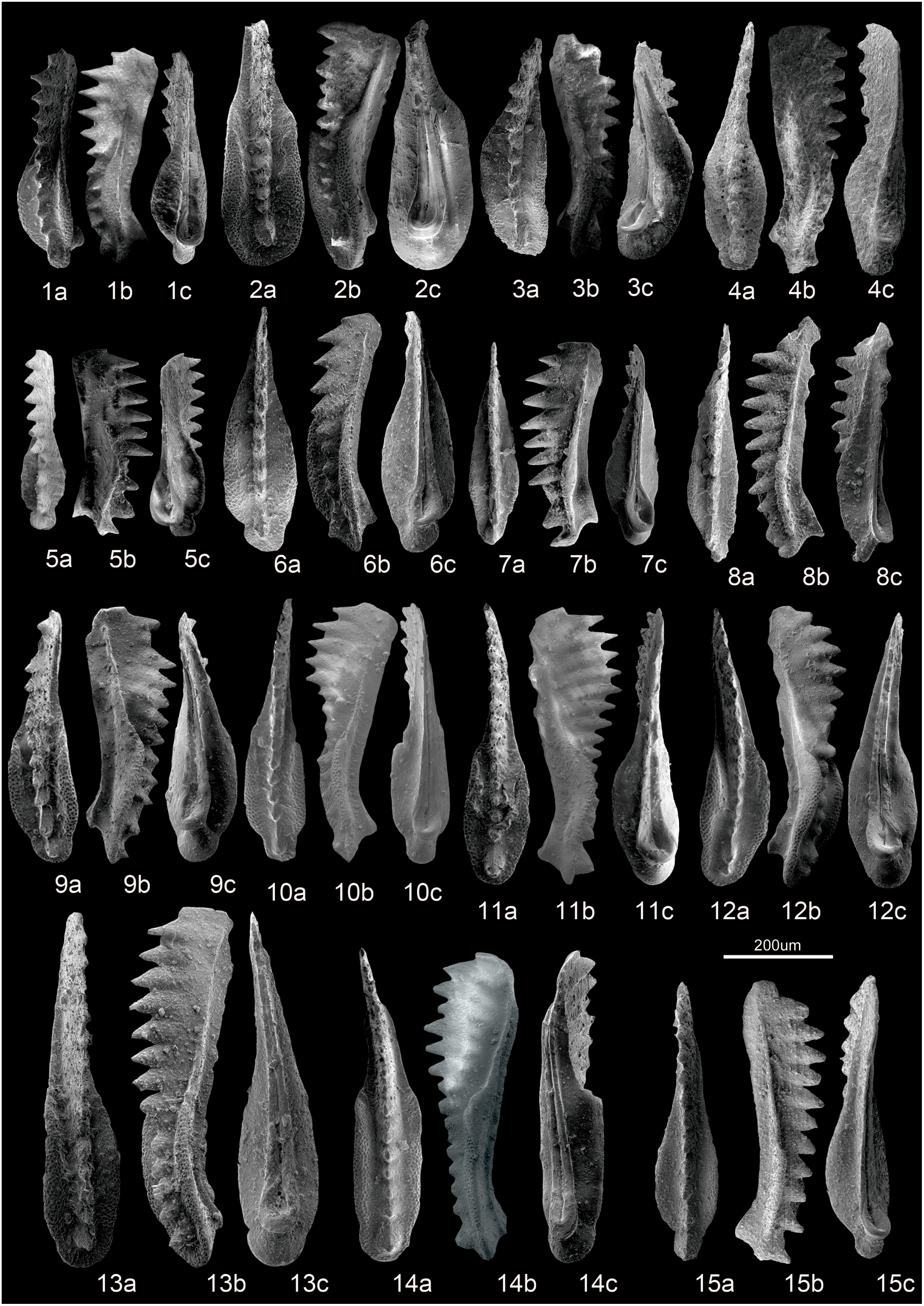
Fig. 7. SEM photographs of conodonts from Ladinian to Carnian strata at Yize. Scale bar = 200 μm. a – upper view; b – for lateral view; c – for lower view. 1 – Q. sp., YZ-88-1-50; 2 – Q. polygnathiformis (Budurov & Stefanov, Reference Budurov and Stefanov1965), YZ-79-022; 3 – P. inclinata (Kovács, Reference Kovács1983), YZ-79-023; 4 – Q. tadpole (Hayashi, Reference Hayashi1968), YZ-79-003; 5 – Q. sp., YZ-46-008; 6 – Q. lobata (Orchard, Reference Orchard, Lucas and Spielmann2007), YZ-12-1-020; 7 – Q. praelindae (Kozur, Reference Kozur2003), YZ-74-014; 8 – Q. praelindae (Kozur, Reference Kozur2003), YZ-74-015; 9 – Q. acuminata (Orchard, Reference Orchard, Lucas and Spielmann2007), YZ-12-1-001; 10 – Q. praelindae (Kozur, Reference Kozur2003), YZ-88-1-58; 11 – Q. tadpole (Hayashi, Reference Hayashi1968), YZ-88-1-47; 12 – Q. intermedia (Orchard, Reference Orchard, Lucas and Spielmann2007), YZ-88-1-42; 13 – P. inclinata (Kovács, Reference Kovács1983), YZ-95-1-020; 14 – K. praeangustus (Kozur, Miräuta & Mock in Kozur, Reference Kozur1980 b), YZ-88-1-39; 15 – Q. praelindae (Kozur, Reference Kozur2003), YZ-95-1-018.
The genus Gladigondolella has a long range from the Spathian to the end of the Julian. A total of five species, including G. arcuata, G. budurovi, G. carinata, G. malayensis and G. tethydis are attributed to this genus (Chen et al. Reference Chen, Krystyn, Orchard, Lai and Richoz2015). G. tethydis is known from the Tethys and Panthalassa, ranging from the Aegean to the uppermost Julian (Igo, Reference Igo1989; Kozur, Reference Kozur1989 a; Gallet et al. Reference Gallet, Besse, Krystyn, Théveniaut and Marcoux1994; Chen et al. Reference Chen, Krystyn, Orchard, Lai and Richoz2015; Kiliç et al. Reference Kiliç, Plasencia, Guex, Hirsch and Montenari2017; Zhang et al. Reference Zhang, Sun, Lai, Joachimski and Wignall2017). G. malayensis is characterized by its broader platform and posteriorly located basal cavity and ranges from the Longobardian to the uppermost Julian (Kozur, Reference Kozur2003; Chen et al. Reference Chen, Krystyn, Orchard, Lai and Richoz2015). G. malayensis and G. tethydis have been reported in Guizhou Province of South China, ranging from the Aegean to the Julian (Lehrmann et al. Reference Lehrmann, Stepchinski, Altiner, Orchard, Montgomery, Enos, Ellwood, Bowring, Ramezani and Wang2015; Sun, Y. D. et al. Reference Sun, Wignall, Joachimski, Bond, Grasby, Lai, Wang, Zhang and Sun2016; Zhang et al. Reference Zhang, Sun, Lai, Joachimski and Wignall2017).
Q. zonneveldi was established in the Julian in British Columbia (Orchard, Reference Orchard, Lucas and Spielmann2007). The reported Q. aff. zonneveldi shares a similar platform outline with Q. zonneveldi but has a sunken carina (Fig. 8).
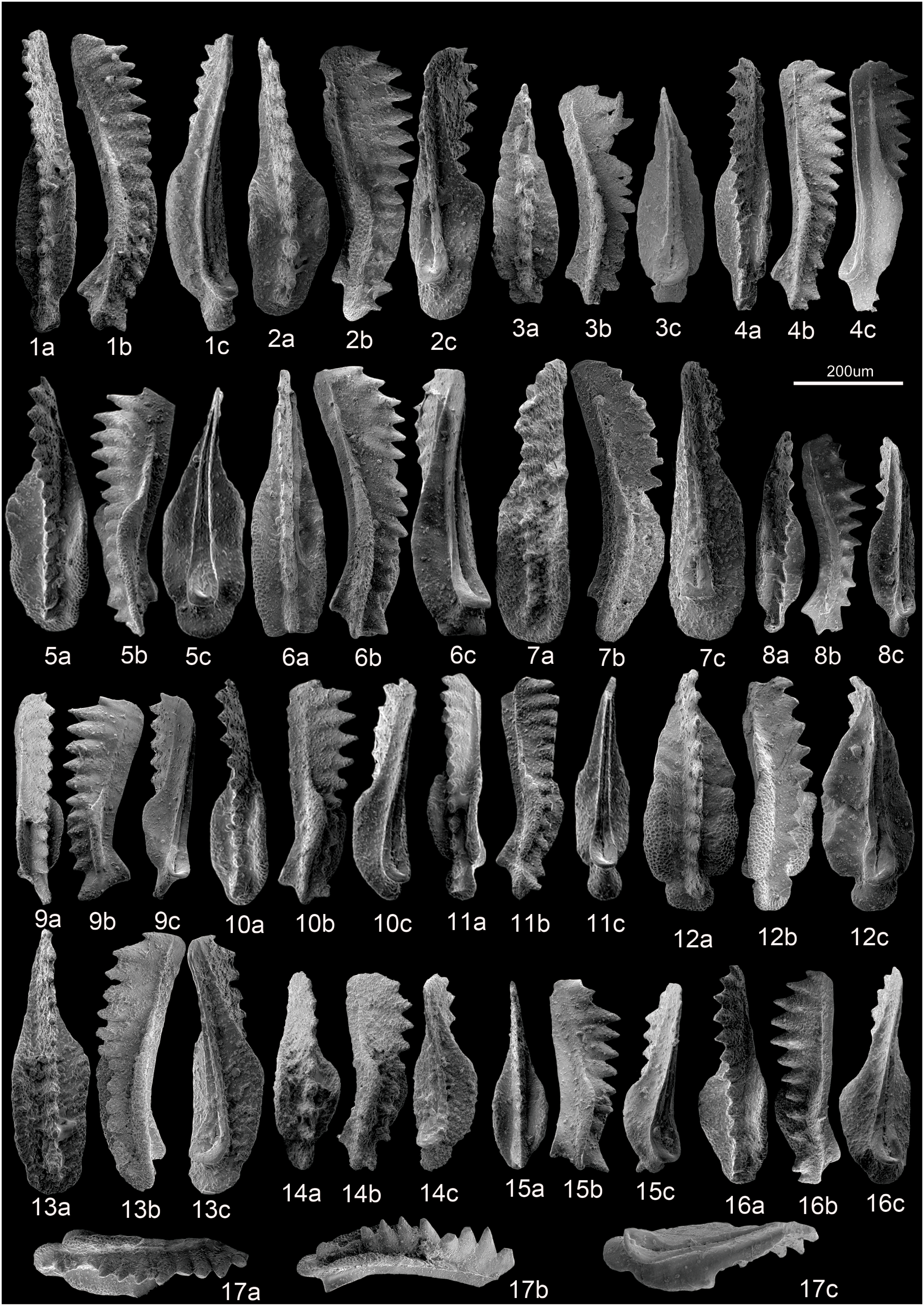
Fig. 8. SEM photographs of conodonts from Ladinian to Carnian strata at Yize. Scale bar = 200 μm. a – upper view; b – lateral view; c – lower view. 1 – Q. praelindae (Kozur, Reference Kozur2003), YZ-95-1-025; 2 – Q. aff. zonneveldi (Orchard, Reference Orchard, Lucas and Spielmann2007), YZ-95-1-049; 3 – Q. lobata (Orchard, Reference Orchard, Lucas and Spielmann2007), YZ-39-1-038; 4 – Q. praelindae (Kozur, Reference Kozur2003), YZ-95-2-032; 5 – Q. lobata (Orchard, Reference Orchard, Lucas and Spielmann2007), YZ-88-1-03; 6 – P. inclinata (Kovács, Reference Kovács1983), YZ-15-001; 7 – Q. polygnathiformis (Budurov & Stefanov, Reference Budurov and Stefanov1965), YZ-39-1-052; 8 – Q. praelindae (Kozur, Reference Kozur2003), YZ-95-2-027; 9 – Q. auriformis (Kovács, Reference Kovács1977), YZ-95-2-038; 10 – Q. tadpole (Hayashi, Reference Hayashi1968), YZ-88-1-06; 11 – Q. praelindae (Kozur, Reference Kozur2003), YZ-88-1-24; 12 – Q. lobata (Orchard, Reference Orchard, Lucas and Spielmann2007), YZ-39-1-009; 13 – P. inclinata (Kovács, Reference Kovács1983), YZ-39-1-055; 14 – Q. auriformis (Kovács, Reference Kovács1977), YZ-99-031; 15 – Q. sp., YZ-12-2-022; 16 – Q. auriformis (Kovács, Reference Kovács1977), YZ-88-1-33; 17 – Q. lobata (Orchard, Reference Orchard, Lucas and Spielmann2007), YZ-39-1-012.
K. praeangustus is the first reported occurrence in southwestern China. The holotype of K. praeangustus was established from Julian strata in Romania, and the species is also known in southern Turkey (Kozur, Reference Kozur1980 a; Chen & Lukeneder, Reference Chen and Lukeneder2017). Compared to Q. polygnathiformis, K. praeangustus has a slender platform with one/two side constriction(s) in the posterior platform. Q. shijiangjunensis and Q. wayaoensis have not yet been reported outside of southwestern China.
4.d. Quadralella auriformis Zone
Lower limit: FO of Q. auriformis.
Upper limit: FO of Q. robusta.
Associated taxa: K. praeangustus, Ma. baloghi, P. inclinata, Q. intermedia, Q. lobata, Q. polygnathiformis, Q. praelindae, Q. spp., Q. tadpole and Q. aff. zonneveldi.
The Q. auriformis Zone ranges from the 67 m to the 72.2 m level. This zone has been established in southwestern China, the Alps and the Himalayas (Gallet et al. Reference Gallet, Besse, Krystyn, Théveniaut and Marcoux1994; Krystyn et al. Reference Krystyn, Balini and Nicora2004; Sun, Reference Sun2006; Hornung et al. Reference Hornung, Brandner, Krystyn, Joachimski, Keim, Lucas and Spielmann2007). The zonal species is characterized by its ear-like platform and differs from Q. tadpole by its expanded middle platform and shorter length (Kovács, Reference Kovács1977; Mastandrea, Reference Mastandrea1995; Kiliç et al. Reference Kiliç, Plasencia, Ishida and Hirsch2015). It ranges from the lower Julian to the lower Tuvalian and is known in the western Tethys, Japan, the Himalayas, northern Oman and southwestern China (Kovács, Reference Kovács1977; Gallet et al. Reference Gallet, Besse, Krystyn, Théveniaut and Marcoux1994; Mastandrea, Reference Mastandrea1995; Hornung et al. Reference Hornung, Brandner, Krystyn, Joachimski, Keim, Lucas and Spielmann2007; Rigo et al. Reference Rigo, Preto, Roghi, Tateo and Mietto2007; Kiliç et al. Reference Kiliç, Plasencia, Ishida and Hirsch2015; Zhang et al. Reference Zhang, Sun, Lai and Wignall2018 b; Sun et al. Reference Sun, Richoz, Krystyn, Zhang and Joachimski2019; Tomimatsu et al. Reference Tomimatsu, Nozaki, Sato, Takaya, Kimura, Chang, Naraoka, Rigo and Onoue2021). The species could be the direct ancestor of Ma. baloghi and Ma. carnica (Fig. 9).

Fig. 9. SEM photographs of conodonts from Ladinian to Carnian strata at Yize. Scale bar = 200 μm. a – upper view; b – lateral view; c – lower view. 1 – Q. praelindae (Kozur, Reference Kozur2003), YZ-88-2-04; 2 – Q. tadpole (Hayashi, Reference Hayashi1968), YZ-88-2-08; 3 – Q. tadpole (Hayashi, Reference Hayashi1968), YZ-88-2-46; 4 – K. praeangustus (Kozur, Miräuta & Mock in Kozur, Reference Kozur1980 b), YZ-88-2-42; 5 – Q. intermedia (Orchard, Reference Orchard, Lucas and Spielmann2007), YZ-88-2-44; 6 – Q. polygnathiformis (Budurov & Stefanov, Reference Budurov and Stefanov1965), YZ-88-3-07; 7 – Q. praelindae (Kozur, Reference Kozur2003), YZ-88-3-16; 8 – Q. auriformis (Kovács, Reference Kovács1977), YZ-88-3-75; 9 – Q. praelindae (Kozur, Reference Kozur2003), YZ-88-3-17; 10 – Q. sp., YZ-88-2-38; 11 – Q. praelindae (Kozur, Reference Kozur2003), YZ-88-1-64; 12 – Q. sp., YZ-88-1-60; 13 – Q. sp., YZ-88-2-43; 14 – Ma. baloghi (Kovács, Reference Kovács1977), YZ-88-1-63; 15 – Q. auriformis (Kovács, Reference Kovács1977), YZ-88-2-26; 16 – Q. sp., YZ-88-2-12.
Ma. baloghi is reported for the first time here in southwestern China. The species shares many common features with Q. auriformis but has distinct nodes on the anterior platform margins (Kovács, Reference Kovács1977; Mastandrea, Reference Mastandrea1995). It has a short range within the Julian substage and is known in Hungary, Oman and Italy (e.g. Kovács, Reference Kovács1977; Mastandrea, Reference Mastandrea1995; Rigo et al. Reference Rigo, Preto, Roghi, Tateo and Mietto2007; Chen et al. Reference Chen, Krystyn, Orchard, Lai and Richoz2015; Kiliç et al. Reference Kiliç, Plasencia, Ishida and Hirsch2015, Reference Kiliç, Plasencia, Guex, Hirsch and Montenari2017; Sun et al. Reference Sun, Richoz, Krystyn, Zhang and Joachimski2019) (Fig. 10).

Fig. 10. SEM photographs of conodonts from Ladinian to Carnian strata at Yize. Scale bar = 200 μm. a – upper view; b – lateral view; c – lower view. 1 – Q. praelindae (Kozur, Reference Kozur2003), YZ-88-3-24; 2 – Q. praelindae (Kozur, Reference Kozur2003), YZ-88-3-38; 3 – Q. intermedia (Orchard, Reference Orchard, Lucas and Spielmann2007), YZ-88-3-32; 4 – Q. sp., YZ-88-3-35; 5 – Q. tadpole (Hayashi, Reference Hayashi1968), YZ-88-3-41; 6 – Q. praelindae (Kozur, Reference Kozur2003), YZ-88-3-43; 7 – Q. auriformis (Kovács, Reference Kovács1977), YZ-88-3-44; 8 – Q. praelindae (Kozur, Reference Kozur2003), YZ-88-3-49; 9 – Q. auriformis (Kovács, Reference Kovács1977), YZ-88-3-60; 10 – Q. tadpole (Hayashi, Reference Hayashi1968), YZ-88-3-61; 11 – Q. tadpole (Hayashi, Reference Hayashi1968), YZ-88-3-63; 12 – Q. tadpole (Hayashi, Reference Hayashi1968), YZ-88-3-66; 13 – Q. auriformis (Kovács, Reference Kovács1977), YZ-88-2-54; 14 – Q. sp., YZ-88-2-63; 15 – Q. tadpole (Hayashi, Reference Hayashi1968), YZ-88-3-05; 16 – Q. auriformis (Kovács, Reference Kovács1977), YZ-88-3-13.
4.e. Quadralella robusta Zone
Lower limit: FO of Q. robusta.
Upper limit: not defined.
Associated taxa: K. praeangustus, P. foliata, P. inclinata, Q. acuminata, Q. auriformis, Q. intermedia, Q. jiangyouensis, Q. lobata, Q. polygnathiformis, Q. praelindae, Q. spp., Q. tadpole and Q. aff. zonneveldi.
The Q. robusta Zone ranges from the 72.2 m level to the top of the section. This regionally important zone was widely recognized in Julian strata in southwestern China (Zhang et al. Reference Zhang, Sun, Lai, Joachimski and Wignall2017, Reference Zhang, Sun, Lai and Wignall2018 b). Q. robusta is characterized by having a large platform and a robust main cusp (in mature form). The species is common in the Zhuganpo and Wayao boundary beds and ranges from the uppermost Julian 1 to the lowermost Tuvalian 1? (Sun, Y. D. et al. Reference Sun, Wignall, Joachimski, Bond, Grasby, Lai, Wang, Zhang and Sun2016; Zhang et al. Reference Zhang, Sun, Lai, Joachimski and Wignall2017, Reference Zhang, Sun, Lai and Wignall2018 b).
5. Discussion
5.a. Conodont palaeogeography and ecology
Early Carnian conodonts show significant provincialism on the globe (Mosher, Reference Mosher1968; Budurov et al. Reference Budurov, Gupta, Sudar and Buryi1985; Lai & Mei, Reference Lai and Mei2000; Klets, Reference Klets2008; Chen et al. Reference Chen, Krystyn, Orchard, Lai and Richoz2015; Martínez-Pérez et al. Reference Martínez-Pérez, Cascales-Miñana, Plasencia and Botella2015; Zhang et al. Reference Zhang, Sun, Lai and Wignall2018 b). The genera Paragondolella, Gladigondolella, Pseudofurnishius, Quadralella, Mazzaella and Budurovignathus dominated the Julian assemblages in the western Tethys, while Mosherella, Paragondolella and Quadralella are common in North America. Only a few monotonous Budurovignathus and a single Gladigondolella species have been reported in North America (e.g. Mosher, Reference Mosher1968; Orchard et al. Reference Orchard, Cordey, Rui, Bamber, Mamet, Struik, Sano and Taylor2001; Balini et al. Reference Balini, Jenks, McRoberts, Orchard, Lucas and Spielmann2007; Orchard & Balini, Reference Orchard, Balini, Lucas and Spielmann2007; Orchard, Reference Orchard and Lucas2010). Mosherella was rare in the western Tethys and only reported in southern Turkey (e.g. Orchard, Reference Orchard and Lucas2010; Chen & Lukeneder, Reference Chen and Lukeneder2017). In the Panthalassa and the Himalayas, Paragondolella, Quadralella, Gladigondolella and Budurovignathus were common in the Julian (e.g. Koike et al. Reference Koike, Kodachi, Matsuno and Baba1991; Krystyn et al. Reference Krystyn, Balini and Nicora2004; Zhang et al. Reference Zhang, Orchard, Algeo, Chen, Lyu, Zhao, Kaiho, Ma and Liu2019).
In South China, conodont assemblages are dominated by genera of Paragondolella and Quadralella in the Julian, while Mosherella, Pseudofurnishius, Gladigondolella and Budurovignathus are very rare (Yang et al. Reference Yang, Liu and Zhang1995, Reference Yang, Hao and Jiang2002; Wang et al. Reference Wang, Wang, Li and Wei2005; Dong & Wang, Reference Dong and Wang2006; Sun, Reference Sun2006; Sun, Y. D. et al. Reference Sun, Wignall, Joachimski, Bond, Grasby, Lai, Wang, Zhang and Sun2016; Sun, Z. Y. et al. Reference Sun, Wignall, Joachimski, Bond, Grasby, Lai, Wang, Zhang and Sun2016; Zhang et al. Reference Zhang, Sun, Lai, Joachimski and Wignall2017, Reference Zhang, Sun, Lai and Wignall2018 b). At the species level, indigenous species are more common. However, Julian conodonts from southwestern China still share many common features with those from the western Tethys and North America, laying a biostratigraphic foundation for supra-regional correlation.
Conodont distribution is controlled, in part, by their ecology, i.e. the dwelling habitats of conodont animals (Kozur, Reference Kozur and Barnes1976; Lai et al. Reference Lai, Wignall and Zhang2001; Chen et al. Reference Chen, Joachimski, Richoz, Krystyn, Aljinović, Smirčić, Kolar-Jurkovšek, Lai and Zhang2021). This is more conspicuous in the Middle and Late Triassic amongst the genera Gladigondolella and Budurovignathus. Gladigondolella was considered a pelagic conodont element restricted to the Tethys realm, part of Panthalassa and the western margin of North America (Kozur, Reference Kozur and Barnes1976; Kozur et al. Reference Kozur, Moix and Ozsvart2009). Two species, namely G. tethydis and G. malayensis, are most common in the western Tethys and Panthalassa but are rather rare in South China (e.g. Koike et al. Reference Koike, Kodachi, Matsuno and Baba1991; Gallet et al. Reference Gallet, Besse, Krystyn, Théveniaut and Marcoux1994; Balini et al. Reference Balini, Germani, Nicora and Rizzi2000; Wang et al. Reference Wang, Wang, Li and Wei2005; Lein et al. Reference Lein, Krystyn, Sylvain and Henry2012; Zhang et al. Reference Zhang, Sun, Lai, Joachimski and Wignall2017; Tomimatsu et al. Reference Tomimatsu, Nozaki, Sato, Takaya, Kimura, Chang, Naraoka, Rigo and Onoue2021). They have never occurred in a platform setting, suggesting the taxa may have favoured deeper water habitats (also see Trotter et al. Reference Trotter, Williams, Nicora, Mazza and Rigo2015).
Abundant and diverse Budurovignathus species occur in various environments in the western Tethys (e.g. Gallet et al. Reference Gallet, Besse, Krystyn, Théveniaut and Marcoux1994; Mastandrea et al. Reference Mastandrea, Neri and Russo1997; Loriga et al. Reference Loriga, Cirilli, Zanche, Bari, Gianolla, Laghi, Lowrie, Manfrin, Mastandrea, Mietto, Muttoni, Neri, Posenato, Rechichi, Rettori and Roghi1998; Balini et al. Reference Balini, Germani, Nicora and Rizzi2000; Mietto et al. Reference Mietto, Andreetta, Broglio, Buratti, Cirilli, De Zanche, Furin, Gianolla, Manfrin, Muttoni, Neri, Nicora, Posenato, Preto, Rigo, Roghi and Spötl2007, Reference Mietto, Manfrin, Preto, Rigo, Roghi, Furin, Gianolla, Posenato, Muttoni and Nicora2012; Richoz et al. Reference Richoz, Krystyn and Spötl2007; Rigo et al. Reference Rigo, Preto, Roghi, Tateo and Mietto2007; Lein et al. Reference Lein, Krystyn, Sylvain and Henry2012; Stocker et al. Reference Stockar, Adatte, Baumgartner and Föllmi2013; Muttoni et al. Reference Muttoni, Mazza, Mosher, Katz, Kent and Balini2014; Karádi et al. Reference Karádi, Budai, Haas, Voros, Piros, Dunkl and Toth2022). Owing to common occurrences of Budurovignathus species in European sections, the FAD of B. diebeli and the last appearance of B. mungoensis were proposed as markers for the L-CB (Kozur & Mostler, Reference Kozur and Mostler1971; Kozur, Reference Kozur1989 b). However, these species are rare in other regions, and the FAD of B. diebeli was later proved in the Longobardian (e.g. Krystyn, Reference Krystyn and Zapfe1983; Loriga et al. Reference Loriga, Cirilli, Zanche, Bari, Gianolla, Laghi, Lowrie, Manfrin, Mastandrea, Mietto, Muttoni, Neri, Posenato, Rechichi, Rettori and Roghi1998; Karádi et al. Reference Karádi, Budai, Haas, Voros, Piros, Dunkl and Toth2022). In South China, Budurovignathus species are very rare and by far best known from Guandao, which represents an oxygenated platform margin environment (Wang et al. Reference Wang, Wang, Li and Wei2005; Enos et al. Reference Enos, Lehrmann, Wei, Yu, Xiao, Chaikin, Minzoni, Berry and Montgomery2006; Lehrmann et al. Reference Lehrmann, Stepchinski, Altiner, Orchard, Montgomery, Enos, Ellwood, Bowring, Ramezani and Wang2015). Budurovignathus specimens are also reported at Guanyinya in northern Sichuan and Dapingzi in Guizhou (Jiang et al. Reference Jiang, Chen and Liu2018, Reference Jiang, Yuan, Chen, Ogg and Yan2019). However, these elements do not develop nodes on their anterior platform margins or have denticles on the platform and are very unusual. In British Columbia and Nevada, Budurovignathus species are found in diverse environments from slope to basin but were never very diverse (Balini & Jenks, Reference Balini, Jenks, Lucas and Spielmann2007; Balini et al. Reference Balini, Jenks, McRoberts, Orchard, Lucas and Spielmann2007; Orchard, Reference Orchard, Lucas and Spielmann2007, Reference Orchard and Lucas2010; Orchard & Balini, Reference Orchard, Balini, Lucas and Spielmann2007). In the Indian Himalayas, Budurovignathus faunas have been obtained from the Kaga and Chomule formations, which represent a neritic shelf setting below the storm wave base (Bhargava et al. Reference Bhargava, Krystyn, Balini, Lein and Nicora2004; Krystyn et al. Reference Krystyn, Balini and Nicora2004; Sun et al. Reference Sun, Richoz, Krystyn, Grasby, Chen, Banerjee and Joachimski2021). Deep-sea sediments in the Panthalassa also contain Budurovignathus faunas (Nakada et al. Reference Nakada, Ogawa, Suzuki, Takahashi and Takahashi2014; Zhang et al. Reference Zhang, Orchard, Algeo, Chen, Lyu, Zhao, Kaiho, Ma and Liu2019; Tomimatsu et al. Reference Tomimatsu, Nozaki, Sato, Takaya, Kimura, Chang, Naraoka, Rigo and Onoue2021). These may indicate that Budurovignathus could thrive in diverse environments. However, why Budurovignathus is so rare in South China remains a mystery.
5.b. The Ladinian–Carnian boundary
Ammonoids and conodonts are favoured as standard biotic indices to define Triassic stage and substage boundaries (Gradstein et al. Reference Gradstein, Ogg, Schmitz and Ogg2020). Daxatina canadensis is a popular choice for the base of the Carnian in the western Tethys, North America and Himalayas (e.g. Krystyn et al. Reference Krystyn, Balini and Nicora2004; Orchard, Reference Orchard, Lucas and Spielmann2007; Mietto et al. Reference Mietto, Manfrin, Preto, Rigo, Roghi, Furin, Gianolla, Posenato, Muttoni and Nicora2012). In South China, ammonoid biostratigraphy is not well established for the Upper Triassic. This is mostly attributed to the paucity of ammonoids commonly found in other regions (e.g. Hsu & Chen, Reference Hsu and Chen1944; Wang, Reference Wang1983; Xu et al. Reference Xu, Niu and Chen2003; Balini et al. Reference Balini, Lucas, Jenks, Spielmann and Lucas2010; Zou et al. Reference Zou, Balini, Jiang, Tintori, Sun and Sun2015; Tong et al. Reference Tong, Chu, Liang, Shu, Song, Song, Song and Wu2019). Some studies implied that the L-CB should be placed in the upper part of the Zhuganpo Fm or even in the lower Wayao Fm (Xu et al. Reference Xu, Niu and Chen2003; Li et al. Reference Li, Sun, Jiang and Hao2013; Zou et al. Reference Zou, Balini, Jiang, Tintori, Sun and Sun2015).
Conodonts are preferred as important markers to define the L-CB. The FAD of several conodont species, including B. diebeli, Mosherella newpassensis and Q. polygnathiformis have been proposed as auxiliary biotic markers for the L-CB (Mosher, Reference Mosher1968; Kozur & Mostler, Reference Kozur and Mostler1971; Mietto et al. Reference Mietto, Andreetta, Broglio, Buratti, Cirilli, De Zanche, Furin, Gianolla, Manfrin, Muttoni, Neri, Nicora, Posenato, Preto, Rigo, Roghi and Spötl2007). However, the FAD of B. diebeli is lower than the FAD of Daxatina canadensis, and B. diebeli is not widely distributed (e.g. Krystyn, Reference Krystyn and Zapfe1983; Loriga et al. Reference Loriga, Cirilli, Zanche, Bari, Gianolla, Laghi, Lowrie, Manfrin, Mastandrea, Mietto, Muttoni, Neri, Posenato, Rechichi, Rettori and Roghi1998; Krystyn et al. Reference Krystyn, Balini and Nicora2004; Mietto et al. Reference Mietto, Andreetta, Broglio, Buratti, Cirilli, De Zanche, Furin, Gianolla, Manfrin, Muttoni, Neri, Nicora, Posenato, Preto, Rigo, Roghi and Spötl2007, Reference Mietto, Manfrin, Preto, Rigo, Roghi, Furin, Gianolla, Posenato, Muttoni and Nicora2012; Orchard, Reference Orchard and Lucas2010; Plasencia et al. Reference Plasencia, Kiliç, Baud, Sudar and Hirsch2018; Karádi et al. Reference Karádi, Budai, Haas, Voros, Piros, Dunkl and Toth2022). Mosherella newpassensis is known in North America and South China but not in the western Tethys (Orchard, Reference Orchard and Lucas2010). The FO of Mosherella newpassensis also falls in the Longobardian (Wang et al. Reference Wang, Wang, Li and Wei2005; Lehrmann et al. Reference Lehrmann, Stepchinski, Altiner, Orchard, Montgomery, Enos, Ellwood, Bowring, Ramezani and Wang2015). The FAD of Q. polygnathiformis is the most suitable as the practical marker for the L-CB, owing to its global distribution and nearly co-occurrence with Daxatina canadensis (Krystyn et al. Reference Krystyn, Balini and Nicora2004; Sun et al. Reference Sun, Hao and Jiang2005; Rigo et al. Reference Rigo, Preto, Roghi, Tateo and Mietto2007; Mietto et al. Reference Mietto, Andreetta, Broglio, Buratti, Cirilli, De Zanche, Furin, Gianolla, Manfrin, Muttoni, Neri, Nicora, Posenato, Preto, Rigo, Roghi and Spötl2007, Reference Mietto, Manfrin, Preto, Rigo, Roghi, Furin, Gianolla, Posenato, Muttoni and Nicora2012; Lehrmann et al. Reference Lehrmann, Stepchinski, Altiner, Orchard, Montgomery, Enos, Ellwood, Bowring, Ramezani and Wang2015). However, Orchard (Reference Orchard and Lucas2010) suggested that the occurrence of Q. polygnathiformis could be affected by variations in sedimentation rates.
In southwestern China, the position of the L-CB is a matter of debate owing to the difficulties in defining the FO of Q. polygnathiformis and a lack of other auxiliary markers. Yang et al. (Reference Yang, Liu and Zhang1995) placed the L-CB in the lowermost Zhuganpo Fm based on massive occurrences of Q. polygnathiformis. Sun et al. (Reference Sun, Hao and Jiang2005) argued that the L-CB should be put at the 3.38 m level in the Zhuganpo Fm based on the evolutionary lineage of P. inclinata – Q. polygnathiformis – Q. maantangensis and the FO of Q. polygnathiformis. Zhang et al. (Reference Zhang, Sun, Lai, Joachimski and Wignall2017) suggested that the L-CB is unlikely lower than the lithological boundary between the Yangliujing and Zhuganpo formations. These minor discrepancies could be due to a lack of robust biostratigraphic control in the L-CB beds and might also point to the diachronous nature of the Zhuganpo Fm.
The Q. intermedia Zone has been proposed as the first Carnian conodont zone in British Columbia. The FO of Q. intermedia is a practical auxiliary biotic marker for the L-CB because it co-occurred with Daxatina canadensis in the Frankites sutherlandi Zone (Balini et al. Reference Balini, Jenks, McRoberts, Orchard, Lucas and Spielmann2007; Orchard, Reference Orchard, Lucas and Spielmann2007, Reference Orchard and Lucas2010; Chen et al. Reference Chen, Krystyn, Orchard, Lai and Richoz2015). At Yize, the FOs of Q. polygnathiformis and Q. intermedia occur at the same level, suggesting that the L-CB should be placed in the lower cherty limestone member. However, age-diagnostic conodonts are lacking from the 14 m to the 29 m level at the transition from the fine-laminated member to the cherty limestone member. It is possible that the FAD of polygnathiformis and intermedia, i.e. the true L-CB, is within this interval.
5.c. Strata correlation using conodont zonation
Short-ranged and widely distributed conodont taxa have been favoured as zonal species. At Yize, the P. inclinata Zone is likely the last Ladinian zone or straddles the L-CB, as the species is known worldwide in various environments (e.g. Kovács, Reference Kovács1983; Orchard, Reference Orchard, Lucas and Spielmann2007; Rigo et al. Reference Rigo, Preto, Roghi, Tateo and Mietto2007; Lein et al. Reference Lein, Krystyn, Sylvain and Henry2012; Lehrmann et al. Reference Lehrmann, Stepchinski, Altiner, Orchard, Montgomery, Enos, Ellwood, Bowring, Ramezani and Wang2015; Tomimatsu et al. Reference Tomimatsu, Nozaki, Sato, Takaya, Kimura, Chang, Naraoka, Rigo and Onoue2021). The established Q. polygnathiformis Zone at Yize could correlate to the Q. intermedia Zone in British Columbia and Nevada and the Q. polygnathiformis IZ in the western Tethys and Panthalassa Ocean (Koike, Reference Koike, Koike and Igo1979; Orchard, Reference Orchard, Lucas and Spielmann2007, Reference Orchard and Lucas2010; Rigo et al. Reference Rigo, Mazza, Karádi, Nicora and Tanner2018). The Q. auriformis Zone is an important zone that is well correlated to the western Tethys, the Indian Himalayas and the Panthalassa due to its clear evolutionary lineage, widespreadness and short range (e.g. Gallet et al. Reference Gallet, Besse, Krystyn, Théveniaut and Marcoux1994; Hornung et al. Reference Hornung, Brandner, Krystyn, Joachimski, Keim, Lucas and Spielmann2007; Tomimatsu et al. Reference Tomimatsu, Nozaki, Sato, Takaya, Kimura, Chang, Naraoka, Rigo and Onoue2021). The Q. robusta Zone could be a practical zone for regional correlation in southwestern China. This zone is immediately below the Ma. carnica Zone and correlates well with the widespread Carnian Humid Episode in the Julian 1 to Julian 2 transition (Hornung et al. Reference Hornung, Brandner, Krystyn, Joachimski, Keim, Lucas and Spielmann2007; Zhang et al. Reference Zhang, Sun and Lai2018 a,b).
5.d. Implications for conodont evolution
Conodonts were reasonably diverse in much of the Late Triassic period and show tendencies toward (i) shortening of the platform, (ii) increasing ornaments on the platform and (iii) forward-shifting of the basal cavity (Mazza et al. Reference Mazza, Furin, Spötl and Rigo2010, Reference Mazza, Cau and Rigo2012; Kiliç et al. Reference Kiliç, Plasencia, Ishida and Hirsch2015; Karádi, Reference Karádi2021).
Shortening of the platform is a general trend that has been observed amongst gondolellids since the late Middle Triassic period. Two lineages, P. excelsa – P. inclinata – P. foliata – Q. tadpole and Q. acuminata – Q. intermedia, were recognized by a decreasing anterior platform from the Longobardian to the Julian in Hungary and British Columbia (Kovács, Reference Kovács1983; Orchard, Reference Orchard, Lucas and Spielmann2007; Orchard & Balini, Reference Orchard, Balini, Lucas and Spielmann2007). Such changes were later widely recognized in the western Tethys and southwestern China (Sun et al. Reference Sun, Hao and Jiang2005; Kiliç et al. Reference Kiliç, Plasencia, Ishida and Hirsch2015, Reference Kiliç, Plasencia, Guex, Hirsch and Montenari2017; Zhang et al. Reference Zhang, Sun, Lai, Joachimski and Wignall2017, Reference Zhang, Sun, Lai and Wignall2018 b).
The ornamented anterior platform is a most distinctive feature amongst gondolellids in the late Longobardian to the Julian. A transition from smooth to nodose or denticulated platforms is first seen in the B. japonica – B. mungoensis – B. diebeli lineage and in B. mostleri (e.g. Balini et al. Reference Balini, Germani, Nicora and Rizzi2000; Chen et al. Reference Chen, Krystyn, Orchard, Lai and Richoz2015; Plasencia et al. Reference Plasencia, Kiliç, Baud, Sudar and Hirsch2018). This evolutionary trend is not yet seen in South China and North America owing to the paucity of Budurovignathus elements. However, a similar evolutionary trend is seen in the Q. auriformis – Ma. baloghi – Ma. carnica lineage in the Julian substage (Kovács, Reference Kovács1977; Mastandrea, Reference Mastandrea1995; Kiliç et al. Reference Kiliç, Plasencia, Ishida and Hirsch2015; Sun et al. Reference Sun, Richoz, Krystyn, Zhang and Joachimski2019).
The forward-shifting pit was typically seen amongst late Tuvalian conodonts, especially in Metapolygnathus and Carnepigondolella (e.g. Mazza et al. Reference Mazza, Furin, Spötl and Rigo2010; Kiliç et al. Reference Kiliç, Plasencia, Guex, Hirsch and Montenari2017). However, in southwestern China, this change seems to occur earlier in the Julian, evidenced by the appearances of Q. langdaiensis, Q. maantangensis, Q. wanlanensis and Q. polygnathiformis magna, which all have obviously forward-shifted pits (Zhang et al. Reference Zhang, Sun, Lai, Joachimski and Wignall2017).
6. Conclusions
Conodont taxonomy and biostratigraphy were investigated in detail at Yize, Yunnan, southwestern China. The Yize conodont fauna represents a typical late Ladinian to early Carnian assemblage with both endemic and cosmopolitan forms. The FOs of Q. polygnathiformis and Q. intermedia are practical markers co-defining the L-CB in the cherty limestone member of the Zhuganpo Fm. Five conodont zones, namely, the P. inclinata Zone, the Q. polygnathiformis Zone, the Q. praelindae Zone, the Q. auriformis Zone and the Q. robusta Zone, are established and represent a practical biostratigraphic framework in the region. The Zhuganpo Fm, one of the most essential Upper Triassic units in South China, could be diachronous and brackets at least the uppermost Ladinian to the lower Carnian.
Acknowledgements
This study is financially supported by the Natural Science Foundation of China [grant No. 41830320, 41821001 and 42272022], State Key Laboratory of Palaeobiology and Stratigraphy (Nanjing Institute of Geology and Palaeontology, CAS) [grant No. 203123] and China Geological Survey [grant No. DD20211391]. Reviewers Viktor Karádi and Yixing Du are thanked for their constructive comments. G. Q. Liu (Nanchang), Q. Wu (Nanjing), J. Chen (Qingdao), Y. Z. Huang (Nanjing), L. N. Wang (Shijiazhuang), X. L. Wu (Wuhan) and J. L. Fu (Wuhan) are thanked for field and lab assistance.
Conflict of interest
None.










