



Significant outcomes
-
The present study found an increased abundance of Lactobacillales (order level), Bacilli (class level) and Actinobacteriota (phylum level) in patients with schizophrenia.
-
The abundance of Bacteroidia (class level), Actinobacteriota (phylum level), Bacteroidota (phylum level), Coriobacteriales (order level) and Coriobacteria (class level) was related to the severity of negative symptoms, general functioning and performance of language abilities.
Limitations
-
All patients in our sample were medicated.
-
Lifestyle characteristics, other than dietary habits, were not controlled in this study.
Introduction
Schizophrenia is a complex neurodevelopmental disorder manifesting in multidimensional psychopathology that includes positive symptoms (delusions and hallucinations), negative symptoms (social withdrawal, diminished emotional reactivity, anhedonia, alogia and avolition) and mood symptoms (Owen et al., Reference Owen, Sawa and Mortensen2016). Moreover, individuals with schizophrenia show impairments across several cognitive domains that are also present in subjects at clinically high risk of psychosis and unmedicated individuals with first-episode psychosis (Moustafa et al., Reference Moustafa, Garami, Mahlberg, Golembieski, Keri, Misiak and Frydecka2016). Although current strategies of pharmacological treatment offer acceptable efficacy with respect to positive symptoms, cognitive deficits and negative symptoms are often resistant to available treatments and largely contribute to impairments of social functioning.
There is a general consensus that schizophrenia is a multi-systemic disorder with a number of biological alterations detectable outside the brain (Pillinger et al., Reference Pillinger, D’Ambrosio, McCutcheon and Howes2019). Among them, subclinical inflammation represents one of the most widely replicated phenomena in subjects with schizophrenia. In this population, subclinical inflammation manifests in altered levels of various leukocyte populations (Karpiński et al., Reference Karpiński, Samochowiec, Frydecka, Sąsiadek and Misiak2018), together with elevated blood and cerebrospinal fluid levels of C-reactive protein (CRP) (Fernandes et al., Reference Fernandes, Steiner, Bernstein, Dodd, Pasco, Dean, Nardin, Gonçalves and Berk2016), pro-inflammatory cytokines (Miller et al., Reference Miller, Buckley, Seabolt, Mellor and Kirkpatrick2011), complement cascade components (Laskaris et al., Reference Laskaris, Zalesky, Weickert, Di Biase, Chana, Baune, Bousman, Nelson, McGorry, Everall, Pantelis and Cropley2019) as well as specific and non-specific antibodies (Ezeoke et al., Reference Ezeoke, Mellor, Buckley and Miller2013). Moreover, there is evidence that subclinical inflammation is associated with neurostructural alterations (Pasternak et al., Reference Pasternak, Kubicki and Shenton2016), cognitive impairments (Bora, Reference Bora2019; Misiak et al., Reference Misiak, Stańczykiewicz, Kotowicz, Rybakowski, Samochowiec and Frydecka2018) and poor response to antipsychotic treatment (Mondelli et al., Reference Mondelli, Ciufolini, Belvederi Murri, Bonaccorso, Di Forti, Giordano, Marques, Zunszain, Morgan, Murray, Pariante and Dazzan2015). Also, a recent meta-analysis by Bora (Reference Bora2019) showed that elevated blood levels of CRP are modestly but significantly associated with impairments of global cognition, verbal and working memory, processing speed as well as planning.
Although a great progress in understanding subclinical inflammation in schizophrenia can be observed in recent years, the exact mechanisms underlying this phenomenon remain unclear. It has been proposed that aberrant immune-inflammatory responses in schizophrenia might be attributed to gut microbiota alterations (Samochowiec & Misiak, Reference Samochowiec and Misiak2021). Moreover, gut microbiota might communicate with the central nervous system through interactions with the autonomic nervous system, and the bloodstream mediators related to gut hormones, the hypothalamic-pituitary-adrenal axis and metabolites released by microbial species (e.g., short-chain fatty acids) (Misiak et al., Reference Misiak, Łoniewski, Marlicz, Frydecka, Szulc, Rudzki and Samochowiec2020). Experimental studies show that the faecal microbiota transplantation from unmedicated patients with schizophrenia induces hyperkinetic behaviours, impairs social interactions and leads to various biological alterations (e.g., up-regulation of the kynurenine-kynurenic acid pathway in the brain and bloodstream as well as altered neurotransmission) in the recipient germ-free mice (Zheng et al., Reference Zheng, Zeng, Liu, Chen, Pan, Han, Liu, Cheng, Zhou, Wang, Zhou, Gui, Perry, Wong, Licinio, Wei and Xie2019; Zhu et al., Reference Zhu, Guo, Wang, Ju, Wang, Ma, Sun, Fan, Xie, Yang, Jie, Zhao, Xiao, Yang, Zhang, Liu, Guo, He, Chen, Chen, Gao, Xu, Yang, Wang, Dang, Madsen, Brix, Kristiansen, Jia and Ma2020a, b). A recent systematic review revealed that individuals with schizophrenia show lower levels of bacterial genera that produce short-chain fatty acids as well as higher levels of the bacteria that produce lactic acid and the bacteria associated with glutamate and GABA metabolism (McGuinness et al., Reference McGuinness, Davis, Dawson, Loughman, Collie, O’Hely, Simpson, Green, Marx, Hair, Guest, Mohebbi, Berk, Stupart, Watters and Jacka2022). Although these alterations overlap with those observed in patients with bipolar disorder or major depression, some observations might be specific to individuals with schizophrenia. These include higher abundance of Prevotella as well as lower abundance of Bacteroides, Haemophilus and Streptococcus (findings consistent across at least 20% of studies included in systematic review).
It should be noted that findings form studies in this field are characterised by relatively low replicability. This observation might originate from the effects of potential confounding factors that are rarely controlled. These include, i.e., recruitment of patients at various stages of illness, medication effects, comorbidities and lifestyle characteristics (e.g., dietary habits and substance use). To the best of our knowledge, recent studies on gut microbiota in schizophrenia were mainly based on the U.S. or Chinese populations, with only few studies representing the European population (Schwarz et al., Reference Schwarz, Maukonen, Hyytiäinen, Kieseppä, Orešič, Sabunciyan, Mantere, Saarela, Yolken and Suvisaari2018; Manchia et al., Reference Manchia, Fontana, Panebianco, Paribello, Arzedi, Cossu, Garzilli, Montis, Mura, Pisanu and Congiu2021). Importantly, there is evidence that geography and ethnicity serve as important determinants of gut microbiota composition (Gupta et al., Reference Gupta, Paul and Dutta2017). A lack of methodological homogeneity is also an important reason underlying low replicability of findings (Nguyen et al., Reference Nguyen, Hathaway, Kosciolek, Knight and Jeste2021). Moreover, it has been shown that gut microbiota alterations might be associated with psychopathological symptoms of schizophrenia, including positive, negative and depressive symptoms (Li et al., Reference Li, Zhuo, Huang, Huang, Zhou, Xiong, Li, Liu, Pan, Li, Chen, Li, Xiang, Wu and Wu2020; Nguyen et al., Reference Nguyen, Kosciolek, Maldonado, Daly, Martin, McDonald, Knight and Jeste2019; Zhu et al., Reference Zhu, Zheng, Ali, Xia, Wang, Chenlong Yao, Chen, Yan, Wang, Chen and Zhang2021). Also, only one study investigated as to whether oropharyngeal microbiota alterations are associated with cognitive impairment observed in schizophrenia (Yolken et al., Reference Yolken, Prandovszky, Severance, Hatfield and Dickerson2021). In light of these shortcomings and research gaps, we aimed to compare gut microbiota composition between stable outpatients with schizophrenia and healthy controls (HCs), controlling for the effects of various confounding factors. Moreover, we aimed to investigate as to whether gut microbiota composition is associated with clinical manifestation and cognitive impairment.
Material and methods
Participants
A total of 53 individuals with schizophrenia and 58 HCs were enrolled at two university hospitals in Wroclaw and Szczecin (Poland) as the convenience sample. All participants were non-consanguineous and represented Caucasian ethnicity. The inclusion criteria were (1) age between 18 and 65 years; (2) a diagnosis of schizophrenia according to the DSM-IV criteria, validated using the Operational Criteria for Psychotic Illness (OPCRIT) checklist (McGuffin et al., Reference McGuffin, Farmer and Harvey1991); (3) maintenance of a stable antipsychotic regimen over the period of at least 6 preceding months and (4) symptomatic remission of positive and disorganisation symptoms based on the Positive and Negative Syndrome Scale (PANSS) items (P1 – delusions, P2 – conceptual disorganisation, P3 – hallucinatory behaviour, G5 – mannerisms/posturing and G9 – unusual thought content rated ≤3) (Andreasen et al., Reference Andreasen, Carpenter, Kane, Lasser, Marder and Weinberger2005). The daily dosage of antipsychotics was expressed as chlorpromazine equivalents (CPZeq). In addition, the current use of antidepressants and mood stabilisers was recorded for all patients.
In turn, HCs had never received psychiatric diagnosis or treatment. They reported no family members affected by psychotic and mood disorders in first- and second-degree relatives. All of them were screened for psychiatric disorders using the Mini-International Neuropsychiatric Interview (M.I.N.I.) (Sheehan, Reference Sheehan1998). Recruitment of HCs was performed through advertisements.
Both groups of participants were matched for age, sex and the level of parental education. The latter one was applied as the proxy measure of socio-economic status.
Clinical assessment
The clinical manifestation was recorded using the following scales: (1) the Positive and Negative Syndrome Scale (PANSS) (Kay et al., Reference Kay, Fiszbein and Opler1987); (2) the Social and Occupational Functioning Assessment Scale (Smith et al., Reference Smith, Ehmann, Flynn, MacEwan, Tee, Kopala, Thornton, Schenk and Honer2011) and (3) the Calgary Depression Scale for Schizophrenia (Addington et al., Reference Addington, Addington and Matickatyndale1994). Two items (N5 – difficulty in abstract thinking and N7 – stereotyped thinking) were excluded from the PANSS score of negative symptoms as there is evidence that they measure other psychopathological constructs (N5 – cognitive symptoms and N7 – thought disorganisation) (Galderisi et al., Reference Galderisi, Mucci, Dollfus, Nordentoft, Falkai, Kaiser, Giordano, Vandevelde, Nielsen, Glenthøj, Sabé, Pezzella, Bitter and Gaebel2021). Due to clinical stability, the associations with positive symptoms were not analysed.
The Repeatable Battery for the Assessment of Neuropsychological Status (RBANS) was used to assess cognitive performance (Randolph et al., Reference Randolph, Tierney, Mohr and Chase1998). The RBANS is based on 12 tasks that allow to examine the following domains of cognition: (1) immediate memory (list learning and story memory); (2) visuospatial/constructional abilities (figure copy and line orientation); (3) language (picture naming and semantic fluency); (4) attention (digit span and coding) and (5) delayed memory (list recall, list recognition, story memory and figure recall). Higher RBSANS scores indicate better cognitive performance.
Dietary habits
The Food Frequency Questionnaire 6 (FFQ-6) was administered to record dietary habits (Niedzwiedzka et al., Reference Niedzwiedzka, Wadolowska and Kowalkowska2019). It is composed of 62 items that record self-reported frequency for the consumption of various food products in the preceding 12 months. Each item measures the frequency of consuming specific food products according to a 6-point scale: 1 – ‘never or almost never’; 2 – ‘once a month or less’; 3 – ‘several times a month’; 4 – ‘several times a week’; 5 – ‘daily’ and 6 – ‘several times a day’. In the present study, we conceptualised dietary habits as the adherence to the Mediterranean diet using the aMED score (Hawrysz et al., Reference Hawrysz, Wadolowska, Slowinska, Czerwinska and Golota2020; Krusinska et al., Reference Krusinska, Hawrysz, Wadolowska, Slowinska, Biernacki, Czerwinska and Golota2018). The aMED score can be calculated based on the frequency of consuming the following food products: (1) vegetables; (2) fruits; (3) whole grains; (4) fish; (5) legumes; (6) nuts and seeds; (7) the ratio of vegetable oils to animal fat and (8) red and processed meat. Participants with the intake above (or below, in case of red and processed meat) the median intake among HCs receive 1 point; otherwise, they receive 0 points. The aMED score ranges between 0 and 8, with higher scores corresponding to greater adherence to the Mediterranean diet.
Stool collection and sequencing
Participants received home stool collection kits and detailed instructions on how to collect stool samples. Returned samples were immediately frozen at –80°C before DNA extraction.
Subsequently, DNA was extracted from frozen faecal samples using the QIAamp PowerFecal Pro DNA Kit (Qiagen) and concentration was measured with the fluorometric method (Quantus, Promega). The libraries were prepared using the QIAseq 16S/ITS region panels according to the standard protocol (Qiagen) for the V3V4 region. The QC libraries were created using the High Sensitivity D1000 ScreenTape System on the TapeStation (Agilent Technologies). The fluorometric technique was applied to measure the library’s concentration (Quantus, Promega). Libraries were diluted according to the QIAseq® 16S/ITS Panel Handbook (Qiagen), pooled and sequenced on the MiSeq (Illumina). Paired-end sequencing was performed using the MiSeq Reagent Kits v3 (600 cycles).
Data processing
Raw sequence data quality was checked using the FastQC application (ver. 0.11.8) and the multiQC application (ver. 1.10.1). The 16S rRNA V3 and V4 primers (341F CCTACGGGNGGCWGCAG and 785R GACTACHVGGGTATCTAATCC) were trimmed off all reads using the cutadapt (version 3.5). The 16S rRNA gene sequencing data were processed using the QIIME2 (Bolyen et al., 2019). Sequence denoising and error correction were carried out with the QIIME2 DADA2 plugin (ver. 1.18.0) (Callahan et al., Reference Callahan, McMurdie, Rosen, Han, Johnson and Holmes2016). The forward and reverse sequences were truncated at positions 235 and 220, respectively. The 5’ ends of the forward and reverse sequences were trimmed off at the first six bases. Otherwise, the default DADA2 parameter settings were used which resulted in the reconstruction of 5036 features and associated amplicon sequence variants (ASVs). Low-frequency features (<10) were removed. The ASVs were then classified using the naïve Bayesian classifier trained on the target (V3–V4) SILVA v138 99% reference sequence database. Sequences classified as chloroplasts, mitochondria, Archaea or Eukarya, and those that could not be classified at the phylum level were removed. Resulting taxa were collapsed by their taxonomic assignment into six levels (i.e., species, genus, family, order, class and phylum). The final table of ASVs, after taxonomy-based filtering and including samples with available metadata (n = 111), contained 3770 features with the median frequency (a total feature count per sample) of 40 921 (range: 7367–157 488). The α-alpha diversity measures were calculated at the level of ASVs and the sampling depth of 7367. The β-diversity (the Bray-Curtis distance) was calculated at each taxonomic level using rarefied tables at depths expressed by the minimum frequency per sample. The feature count tables were rarefied using the rtk package (Saary et al., Reference Saary, Forslund, Bork and Hildebrand2017).
Statistical analysis and bioinformatics
Between-group differences in general characteristics were assessed using the χ2 test (categorical variables) and the Mann–Whitney U test (continuous variables). The level of significance in bivariate tests of general characteristics was set at p < 0.05.
Multivariate analysis included the distance-based redundancy analysis (dbRDA) based on the Bray–Curtis dissimilarities using the capscale function, while the selection of significant variables was carried out with the ordistep function from the vegan package (Oksanen et al., Reference Oksanen, Blanchet, Friendly, Kindt, Legendre, McGlinn, Minchin, O.ʼHara, Simpson, Solymos, Stevens, Szoecs and Wagner2020) using the combined (forward and backward) stepwise selection algorithm, only for models that were significant in the global test (when all variables were included). The ordistep function enables automatic model building for constrained ordination methods (such as the dbRDA). The scores from the simplified dbRDA models (with significant explanatory variables) were used to create ordination diagrams using the scaling with the main interest focused on the relationship between bacterial features and explanatory variables. In this scaling, relative direction of arrows approximates the linear correlation between the bacterial features, between the bacterial features and characteristics of participants (explanatory variables) as well as between explanatory variables.
To examine the gut microbiome compositional differences between patients with schizophrenia and HCs, we performed the differential abundance analysis using both univariate and multivariate analyses based on the generalised linear model with a negative binomial distribution and the log-link function. The generalised linear models for multivariate abundance data were fitted using the mvabund package (Wang et al., Reference Wang, Naumann, Wright and Warton2012). The default family, when fitting multivariate model, was negative binomial, assuming a quadratic mean-variance relationship and a log-linear relationship between the dependent and independent variables. A multivariate hypothesis of the difference in the community composition between groups was tested using the analysis of deviance and the log-likelihood ratio test statistic. The family-wise error rates were analysed with the Westfall and Young’s stepdown resampling algorithm (SRA) in the generalised linear model univariate analysis. The relationships between bacterial features and clinical characteristics were assessed using the Spearman correlation coefficients followed by the false discovery rate (FDR) procedure.
Results
The general characteristics of the sample are reported in Table 1. Both groups did not differ significantly in terms of age, sex and the level of parental education. However, significant between-group differences with respect to the level of education were found. Individuals with schizophrenia also presented significantly higher BMI, higher level of nicotine dependence and worse cognitive performance across all domains, except for the RBANS language domain. The frequency of using of concomitant, non-psychiatric medications was similar in both groups.
Table 1. General characteristics of the sample
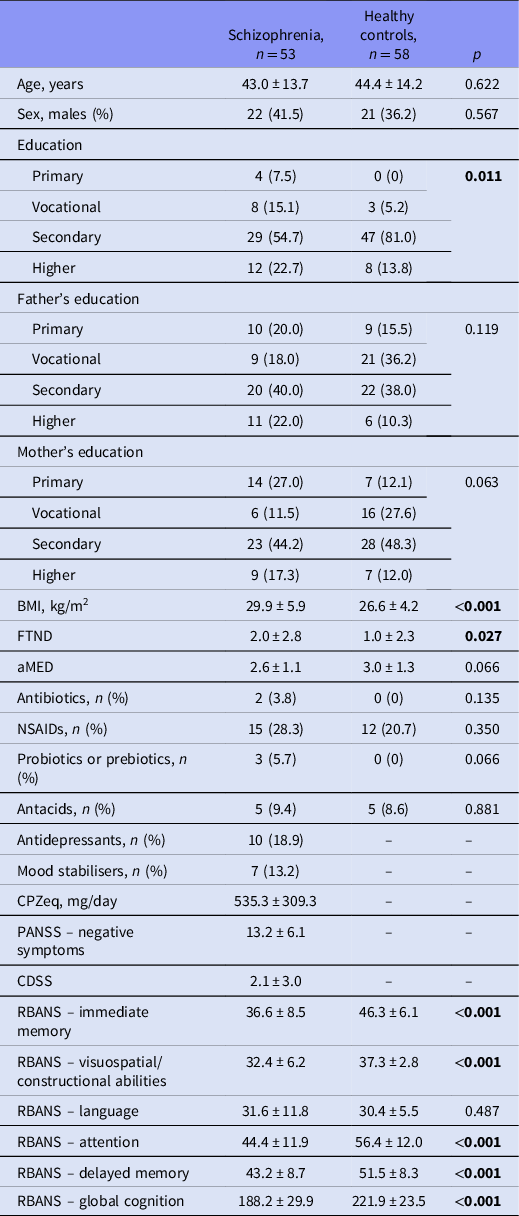
aMED, the Adherence to Mediterranean Diet score; BMI, body mass index; CDSS, the Calgary Depression Scale for Schizophrenia; CPZeq, chlorpromazine equivalent dosage; FTND, the Fagerstrom Test for Nicotine Dependence (Pomerleau et al., Reference Pomerleau, Majchrzak and Pomerleau1989), NSAIDs, non-steroid anti-inflammatory drugs; PANSS, the Positive and Negative Syndrome Scale; RBANS, the Repeatable Battery for the Assessment of Neuropsychological Status.
Significant differences (p < 0.05) were marked with bold characters.
There were no significant between-group differences with respect to α-diversity measures (Fig. 1). The dbRDA was used to assess the effect of patients’ characteristics on the gut microbial community structure (at various taxonomic levels). The first dbRDA analysis was carried out in both subgroups of participants and included the group status (individuals with schizophrenia vs. HCs), age, sex, BMI and the level of education. Explained variance (expressed by adjusted R 2) for simplified models with significant explanatory variables only (selected by the vegan ordistep function) varied from 2.3% for the species to 11.3% for the class. Thus, explanatory power of these models was limited. The summary of the full and simplified models (with significant explanatory variables only), including the adjusted R 2, is presented in Supplementary Table 1. Group status (individuals with schizophrenia vs. HCs) was found to be significantly associated with the gut community composition at the higher taxonomic ranks, i.e., the order, class and phylum (Fig. 2). Triplots of the dbRDA output showed that the taxa that were enriched in subjects with schizophrenia represented three orders (Erysipelotrichales, Bifidobacteriales and Lactobacillales), two classes (Bacilli and Actinobacteria) and one phylum (Actinobacteriota). In turn, the taxa that were enriched in HCs represented the order Bacteroidales and the class Bacteroidia.

Fig. 1. The comparison of α-diversity measures in patients with schizophrenia (SCZ) and healthy controls (HCs).

Fig. 2. The triplot ordination diagrams from the Bray–Curtis distance-based RDA analysis at the level of order (A), class (B) and phylum (C). The analysis includes the effects of age, body mass index (BMI), education and group status (schizophrenia vs. healthy controls). Diagrams show the simplified models limited to significant effects. The triplot shows correlations between gut microbes (red arrows) and explanatory variables (blue arrows). Projection of the group status centroids (blue diamonds) on gut microbe vectors approximates their abundance. Abbreviations: F – females; H – higher education; HCs – healthy controls; M – males; P – primary education; V – vocational education; S – secondary education; SCZ – patients with schizophrenia.
To identify the bacterial features that were differentially abundant in subjects with schizophrenia and HCs, a set of univariate tests were further performed. These tests revealed that the order Lactobacillales, classes Bacilli and Actinobacteria as well as the phylum Actinobacteriota were enriched in subjects with schizophrenia (Fig. 3). Between-group differences in the abundance of Lactobacillales, Bacilli and Actinobacteriota remained significant after including age, sex, BMI and education as covariates in the generalised linear model analysis (Supplementary Tables 2 and 3).

Fig. 3. Results of the differential abundance analysis at the level of order (A), class (B) and phylum (C). The significance of the individual taxa based on the generalised linear model analysis (assuming negative binomial distribution and log-link function) is indicated by the size of the point (triangle) – large triangle indicates the corrected p-value < 0.05. Abbreviations: GLZ – generalised linear model; SRA – Westfall and Young’s stepdown resampling algorithm; SCZ – schizophrenia.
Finally, a series of bivariate correlations between clinical characteristics and bacterial features were tested using the Spearman coefficients and the FDR procedure (Fig. 4). At the order level, a significant negative correlation between the abundance of Coriobacteriales and the RBANS language score was observed. At the class level, significant correlations of the abundance of Bacteroidia with the SOFAS score, the score of PANSS negative subscale and the RBANS language score were found. Moreover, there was a significant negative correlation of the abundance of Coriobacteria with the RBANS language score. Finally, at the phylum level, significant correlations of the abundance of Bacteroidota with the SOFAS score, the PANSS negative symptoms score and the RBANS language score were found. The abundance of Actinobacteriota (phylum level) was significantly and negatively correlated with the RBANS language score. No significant associations with CPZeq as well as the use of antidepressants and mood stabilisers were found.

Fig. 4. Bivariate correlations of bacterial abundancies with clinical characteristics at the level of species (A), genus (B), family (C), order (D), class (E) and phylum (F). Abbreviations: AD – antidepressants; CDSS – the Calgary Depression Scale for Schizophrenia; CPZeq – chlorpromazine equivalent dosage; MS – mood stabilisers; PANSS (N) – the Positive and Negative Syndrome Scale – negative symptoms subscore; RBANS (A) – the Repeatable Battery for the Assessment of Neuropsychological Status – attention score; RBANS (DM) – the Repeatable Battery for the Assessment of Neuropsychological Status – delayed memory subscore; RBANS (IM) – the Repeatable Battery for the Assessment of Neuropsychological Status – immediate memory subscore; RBANS (VC) – the Repeatable Battery for the Assessment of Neuropsychological Status – visuospatial/constructional abilities subscore; SOFAS – the Social and Occupational Functioning Assessment Scale.
Discussion
Main findings of the present study indicate that a number of bacteria at various taxonomic levels might show altered abundance in patients with schizophrenia. However, only some of them might be present regardless of potential confounding factors and the method used to analyse the data. These include enrichment of Lactobacillales (the order level), Bacilli (the class level) and Actinobacteriota (the phylum level). Importantly, none of these observations appeared to be associated with the dosage of antipsychotics or the use of antidepressants and mood stabilisers. Also, these findings cannot be attributed to between-group differences in dietary habits operationalised as the aMED score. Our findings are partially in agreement with previous studies showing greater abundance of the phylum Actinobacteria (Li et al., Reference Li, Zhuo, Huang, Huang, Zhou, Xiong, Li, Liu, Pan, Li, Chen, Li, Xiang, Wu and Wu2020), the family Lactobacillaceae (Ma et al., Reference Ma, Asif, Dai, He, Zheng, Wang, Ren, Tang, Li, Jin, Li and Chen2020) and the genus Lactobacillus (Shen et al., Reference Shen, Xu, Li, Huang, Yuan, Wang, Zhang, Hu and Liang2018) in patients with chronic schizophrenia. In the systematic review, McGuinness et al. (Reference McGuinness, Davis, Dawson, Loughman, Collie, O’Hely, Simpson, Green, Marx, Hair, Guest, Mohebbi, Berk, Stupart, Watters and Jacka2022) found that increased abundance of the Lactobacillus genus is shared by schizophrenia, bipolar disorder and major depression. Therefore, this observation may not be associated with specific severe mental illness. However, it is important to note that Lactobacilli express glutamate decarboxylase that is responsible for the conversion of L-glutamate to GABA (Yogeswara et al., Reference Yogeswara, Maneerat and Haltrich2020). In turn, Actinobacteriota play a number of physiological roles that might be relevant for the pathogenesis of schizophrenia. These include the regulation of glucose homeostasis, interactions with the immune system through the induction of regulatory T cells and the impact on neurotransmission through the involvement in the tryptophan metabolism pathway (Binda et al., Reference Binda, Lopetuso, Rizzatti, Gibiino, Cennamo and Gasbarrini2018; Kaur et al., Reference Kaur, Bose and Mande2019). In parallel, the role of inflammation in the development of schizophrenia seems to be associated with microbiome alterations. Studies have shown that increased abundance of bacteria producing lactic acid, such as the Lactobacillus and Bifidobacterium genus may impact gut inflammation (Xu et al., Reference Xu, Wu, Liang, He, Gu, Li, Luo, Chen, Gao, Wu, Wang, Zhou and Wang2020). Moreover, the Lactobacillus genus might stimulate the production of tumour necrosis factor in patients with schizophrenia (He et al., Reference He, Kosciolek, Tang, Zhou, Li, Ma, Zhu, Yuan, Yuan, Li, Jin, Knight, Tsuang and Chen2018). Finally, various bacteria from the class Bacilli can produce dopamine and noradrenaline, thereby influencing the gut–brain axis (Strandwitz, Reference Strandwitz2018).
The present study also demonstrated a number of correlations between various bacterial taxa (including the Bacteroidia class, the Actinobacteriota phylum, the Bacteroidota phylum, the Coriobacteriales order and the Coriobacteria class) and clinical characteristics (the severity of negative symptoms, performance of the language cognitive domain as well as the level of social and occupational functioning). However, it should be noted that most of them (except for the Actinobacteriota phylum) were not related to differentially abundant taxa. Moreover, although all of these bacterial taxa were related to performance of language abilities, this cognitive domain was not found to be impaired in patients with schizophrenia from our sample. To our knowledge, none of previous studies showed similar patterns of correlations between the abundance of specific bacterial taxa and symptomatic manifestation of schizophrenia. Interestingly, one study found enriched abundance of Bifidobacterium longum genus (Actinobacteriota phylum level) in schizophrenia and its correlation with impaired performance of several cognitive domains (Zhu et al., Reference Zhu, Ju, Wang, Wang, Guo, Ma, Sun, Fan, Xie, Yang, Jie, Zhao, Xiao, Yang, Zhang, Feng, Guo, He, Chen, Chen, Gao, Xu, Yang, Wang, Dang, Madsen, Brix, Kristiansen, Jia and Ma2020b). Moreover, greater abundance of the Lactobacillus group (Lactobacillales order level) was associated with worse global functioning in patients with first-episode psychosis (Schwarz et al., Reference Schwarz, Maukonen, Hyytiäinen, Kieseppä, Orešič, Sabunciyan, Mantere, Saarela, Yolken and Suvisaari2018).
Importantly, this study is characterised by various limitations that need to be considered. Our sample was not large, and thus the likelihood of false positive and false negative findings cannot be excluded. Although both groups of participants were matched for age, sex and the level of parental education as the proxy measure of socio-economic position, the possibility of selection bias should be indicated. For instance, both groups did not differ significantly in terms of dietary habits. However, our previous study showed that poor adherence to Mediterranean diet appears only in patients with the deficit subtype of schizophrenia, characterised by primary and enduring negative symptoms (Kowalski et al., Reference Kowalski, Bogudzińska, Stańczykiewicz, Piotrowski, Bielawski, Samochowiec, Szczygieł, Plichta and Misiak2022). Another limitation is that all patients in our sample were medicated, although we controlled for a number of clinical characteristics related to medication effects. Moreover, certain lifestyle characteristics that might affect gut microbiota were not controlled in this study. These include, i.e., physical activity as well as the use of alcohol and illicit drugs. Deep whole-genome sequencing and metabolomic analysis could provide additional insights into the functional relationship between microbiota and symptoms of schizophrenia. Finally, a lack of longitudinal design does not allow to establish conclusions about causality.
In sum, our study shows altered gut microbiota composition in patients with schizophrenia from Europe. Specifically, our findings indicate increased abundance of Lactobacillales (order level), Bacilli (class level) and Actinobacteriota (phylum level) in this population after controlling for the effects of various confounding factors. Certain gut bacteria (the Bacteroidia class, the Actinobacteriota phylum, the Bacteroidota phylum, the Coriobacteriales order and the Coriobacteria class) might also be related to clinical manifestation of schizophrenia in terms of the severity of negative symptoms, general functioning and performance of language abilities. However, additional studies, adopting longitudinal designs, homogeneous methodology and controlling for latent confounding are needed before their translation into clinical practice.
Supplementary material
To view supplementary material for this article, please visit https://doi.org/10.1017/neu.2022.38
Acknowledgements
Authors are deeply grateful to all patients and HCs participating in this study.
Author contributions
BM – conceptualisation, study design, recruitment of patients and healthy controls, data analysis and manuscript writing; PŻ, DK and ŁŁ – analysis of gut microbiota and manuscript writing; KK and IŁ – manuscript writing; JS, MJ, AC and Patryk Piotrowski – recruitment and psychiatric assessment of patients, critical review of the manuscript; Piotr Plichta – assessment of neurocognition and manuscript editing; MK – bioinformatic analysis
Financial support
This study received funding from the OPUS grant awarded by National Science Centre, Poland (grant number: 2018/31/B/NZ5/00527).
Conflict of interest
I.Ł. is a shareholder of the company involved in probiotic selling and the multiomic analysis of microbiota. Other authors declare no conflict of interest.
Ethical statement
The study was approved by the Bioethics Committees at Wroclaw Medical University (Wroclaw, Poland; approval number: 512/2019) and Pomeranian Medical University (Szczecin, Poland; approval number: KB-0012/130/2019). It was performed in agreement with the Declaration of Helsinki and all participants gave written informed consent.







